
Michael Pollay “The function and structure of the cerebrospinal fluid outflow system“, Cerebrospinal Fluid Research 2010, 7:9.
La presente revisione delinea lo sviluppo della nostra comprensione dell’anatomia e delle proprietà fisiologiche dei due sistemi responsabili del drenaggio del fluido cerebrospinale (FCS) nella circolazione sistemica. I ruoli dei sistemi di deflusso dei villi aracnoidei cranici e spinali (VA) e di quello linfatico sono valutati sulla base della predominanza di uno sull’altro in diverse specie e su più gradi di maturazione animale.
Scarica la sintesi in italiano in formato pdf (2M)
Introduzione
L’anatomia funzionale delle strutture responsabili del ritorno del fluido cerebrospinale (FCS) alla circolazione generale è basata su prove alquanto contrastanti per quanto riguarda la loro ubicazione, le rispettive caratteristiche anatomiche e capacità funzionali. I due sistemi di deflusso per l’uscita del FCS dal compartimento intracranico del FCS controllano l’equilibrio tra la produzione di FCS ed il drenaggio che in ultimo influisce sulla costanza dell’ambiente cerebrale. La nostra comprensione attuale dei due sistemi principali di drenaggio funzionale del FCS si è evoluta separatamente rispetto al riconoscimento del percorso dei VA, cosa che si è verificata molto prima della presa di coscienza dell’importanza della via di deflusso linfatico.
Storia antica dello studio dei villi aracnoidei cranici e spinali
Vesalio e Willis notarono entrambi la presenza delle granulazioni aracnoidee nel XVI e nel XVII secolo. La dissezione dettagliata di Pacchioni nel 1705 rivelò per la prima volta la loro relazione con il seno sagittale, che gli suggerì il ruolo secretorio di tali strutture. Verso la fine del XIX secolo, Luschka notò anch’egli le strutture aracnoidee che penetravano nelle lacune del seno sagittale, cosa che gli suggerì una funzione speciale di tali proiezioni aracnoidee. Egli notò inoltre che le granulazioni aracnoidee rappresentavano un allargamento dei villi normali dell’aracnoide [1]. Trolard confermò tali scoperte nel 1870 quando descrisse le proiezioni sia nelle lacune laterali sia direttamente nel seno sagittale superiore [2]. Quasi al contempo, Quincke seguì la distribuzione del cinabro iniettato nel FCS degli animali ed osservò al microscopio il materiale avviluppato nelle granulazioni aracnoidee; ciò gli suggerì il ruolo dei villi nella rimozione del FCS [3]. Successivamente, Key e Retzius [4] confermarono questa conclusione negli esseri umani mediante l’iniezione di gelatina colorata in campioni di cervello; essi scoprirono che tale sostanza era penetrata nelle strutture dei villi, entrando nelle lacune laterali e nel seno venoso, in modo da far ritornare il FCS nel sangue. È inoltre interessante notare che alcuni dei traccianti colorati (trypan blu) usati nei suoi esperimenti sono stati trovati nei linfonodi cervicali.
Cushing (1901) credette inizialmente che le proiezioni aracnoidee dovessero avere una struttura valvolare per soddisfare i requisiti di trasferimento unidirezionale del FCS nella circolazione sistemica. Fu successivamente convinto da Weed (1914) che tali strutture rappresentavano un diverticolo cieco semipermeabile inframezzato tra il sangue venoso nei seni cerebrali ed il liquor nello spazio subaracnoideo [5]. Si può presumere che sia le differenze osmotiche che quelle idrostatiche siano servite da filtro per il fluido dallo spazio subaracnoideo ai seni venosi.
In una prima revisione, tuttavia, Davson [6] concluse che gli studi fisiologici che dimostravano la rimozione delle materie proteiche e del particolato dallo spazio subaracnoideo ai seni cranici avessero portato a credere che i villi aracnoidei (VA) fossero una tasca cieca semipermeabile insostenibile e che l’unica forza necessaria per ottenere il passaggio unidirezionale del fluido nel sangue venoso fosse una pressione idrostatica differenziale favorevole. Quasi mezzo secolo dopo, la tesi di Weed sulla forma e la funzione dei VA venne confutata e fu proposto un nuovo concetto riguardante l’anatomia funzionale del drenaggio del FCS dallo spazio subaracnoideo cranico [5].
È stato necessario attendere fino al XX secolo prima che il rapporto funzionale tra i VA spinali e le vene della radice spinale ed il sistema linfatico venisse completamente considerato, e ciò nonostante Elman (1923) avesse descritto lo spazio subaracnoideo intorno alla radice dorsale dei nervi spinali che conteneva cluster di cellule aracnoidee [7]. Inoltre, egli osservò la distribuzione dei granuli blu di Prussia, in seguito ad iniezione subaracnoidea, in questi cluster, nella dura spinale regionale e nelle vene spinali della zona. Utilizzando traccianti chimici, Wisclocki [8] descrisse un ricco plesso di vasi intorno alla radice spinale dorsale che trovò simile ai seni venosi intracranici per quanto riguarda la forma e la prossimità allo spazio subaracnoideo spinale. Ciò suggerì un rapporto meningeo con questi canali venosi simile a quello riscontrato nel compartimento cranico. Questa relazione e la funzione potenziale nel drenaggio del FCS furono maggiormente considerate solo in seguito, verso la metà del XX secolo.
Un concetto più nuovo di sistema di drenaggio dei villi aracnoidei
La revisione della tesi precedente sul sistema di deflusso del FCS cranico iniziò con gli studi di Welch e colleghi sui VA isolati ed il concetto di valvole del FCS. Welch e Friedman [5] ritennero che la prima considerazione di Cushing sui VA fosse in effetti corretta.
Ciò si basava sulla perfusione dei VA cranici isolati nei cercopitechi gialloverdi (Figura 1). Dimostrarono con studi al microscopio ottico che i villi consistevano in un labirinto di tubi aperti quando il tessuto era fissato a pressioni fisiologiche normali. Questi tubi aracnoidei venivano cancellati quando il flusso del liquido di fissaggio si trovava nella direzione opposta (Figura 2). I dati fisiologici di tale studio hanno corroborato questa visione anatomica del sistema del FCS con flusso nella direzione del seno che inizia da 20 a 50 mm H20 con una curva convessa rispetto all’asse della pressione (Figura 3). Il flusso del liquido di perfusione in risposta alla pressione non è andato nella direzione opposta, come ci si sarebbe potuti aspettare dalla disposizione anatomica presentata. Welch e Pollay [9] dimostrarono in seguito il flusso di particelle attraverso i VA usando la stessa disposizione sperimentale. Questi studi hanno dimostrato che le particelle di dimensioni micrometriche erano in grado di attraversare l’interfaccia del seno. Inoltre, l’aumento della concentrazione di proteine del perfusato non ha impedito il passaggio del preparato dal lato meningeo a quello del seno, anche se la maggiore viscosità del liquido ha fatto rallentare lievemente la velocità di flusso come previsto. Questa scoperta ha corroborato gli studi precedenti, che hanno dimostrato che l’interfaccia dei VA è permeabile alla materia proteica ed al particolato. Si è inoltre osservato che l’aggiunta di una sostanza con attivazione superficiale (Tween 80) ha abbassato in modo significativo la pressione di apertura necessaria per avviare il flusso dal lato meningeo a quello del seno del preparato sperimentale. Quest’osservazione ha suggerito che la pressione di apertura del passaggio tubolare sia dovuta alla viscosità intrinseca della struttura valvolare.
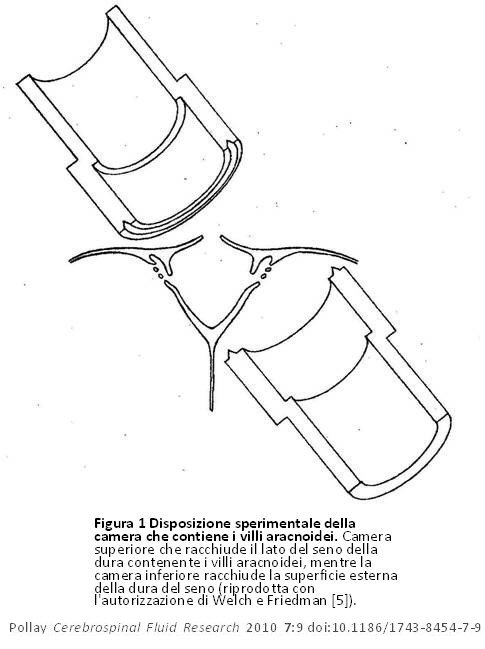 Figura 1 Disposizione sperimentale della camera che contiene i villi aracnoidei. Camera superiore che racchiude il lato del seno della dura contenente i villi aracnoidei, mentre la camera inferiore racchiude la superficie esterna della dura del seno (riprodotta con l’autorizzazione di Welch e Friedman [5]). Figura 1 Disposizione sperimentale della camera che contiene i villi aracnoidei. Camera superiore che racchiude il lato del seno della dura contenente i villi aracnoidei, mentre la camera inferiore racchiude la superficie esterna della dura del seno (riprodotta con l’autorizzazione di Welch e Friedman [5]). |
 Figura 2 Le valvole del flusso cerebrospinale dopo l’inversione del gradiente di pressione dal FCS al sangue. A: Struttura a villo aperto (VO) quando il gradiente di pressione è positivo. B: Villo collassato o chiuso (VC) quando il gradiente è negativo (riprodotto con l’autorizzazione di Welch e Friedman [5]). Figura 2 Le valvole del flusso cerebrospinale dopo l’inversione del gradiente di pressione dal FCS al sangue. A: Struttura a villo aperto (VO) quando il gradiente di pressione è positivo. B: Villo collassato o chiuso (VC) quando il gradiente è negativo (riprodotto con l’autorizzazione di Welch e Friedman [5]). |
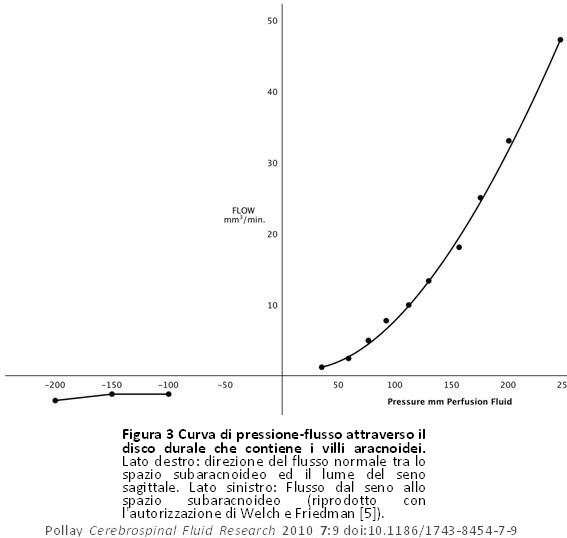 Figura 3 Curva di pressione-flusso attraverso il disco durale che contiene i villi aracnoidei. Lato destro: direzione del flusso normale tra lo spazio subaracnoideo ed il lume del seno sagittale. Lato sinistro: Flusso dal seno allo spazio subaracnoideo (riprodotto con l’autorizzazione di Welch e Friedman [5]). Figura 3 Curva di pressione-flusso attraverso il disco durale che contiene i villi aracnoidei. Lato destro: direzione del flusso normale tra lo spazio subaracnoideo ed il lume del seno sagittale. Lato sinistro: Flusso dal seno allo spazio subaracnoideo (riprodotto con l’autorizzazione di Welch e Friedman [5]). |
Anatomia dello sviluppo dei villi aracnoidei
Si ritiene solitamente che le strutture derivate dal tessuto aracnoideo (villi aracnoidei, VA) siano inizialmente una proiezione di dimensioni microscopiche che non penetra completamente la parete durale del seno venoso cranico. Con la maturazione, continuano a crescere di dimensione e penetrano la dura madre con cambiamenti nella morfologia funzionale di queste strutture, che può quindi essere vista senza ingrandimento. In questa fase sono propriamente definite granulazioni aracnoidee o corpi di Pacchioni. È stato proposto che la maturità e l’aumento di pressione all’interno del sistema del FCS forniscano lo stimolo necessario per questa trasformazione [11].
Nell’uomo, queste strutture sono presenti in gran numero lungo i seni venosi cerebrali, ma soprattutto lungo il seno sagittale superiore nella regione in cui la vena occipito-parietale entra nella cavità del seno venoso. Inoltre, sono visibili con una certa regolarità e densità lungo il seno venoso trasverso, soprattutto nella regione di confluenza dei seni. I villi aracnoidei nella regione della sella turcica sono stati osservati anche negli esseri umani con un seno intracavernoso o un plesso venoso ben sviluppati [12].
Nei feti umani e nei neonati, Gomez et al [13] hanno dimostrato nel feto di 26 settimane, delle depressioni durali nella parete durale venosa. Queste depressioni contenevano cluster di cellule aracnoidee che penetravano tra le fibre durali ed alcune raggiungevano una posizione sub-endoteliale. Entro le 35 settimane, queste depressioni sono state trasformate in semplici protrusioni attraverso la dura con le caratteristiche dei VA. Dalle 39 settimane in poi, il numero e la complessità dei villi sono aumentati. Questi risultati sono generalmente in accordo con studi precedenti meno approfonditi sull’aspetto delle depressioni durali e corrispondono con la maturazione osservata dello spazio subaracnoideo entro la 30a settimana di vita fetale[3]. La relativa scarsità di queste strutture nel tardo periodo fetale e all’inizio di quello postnatale suggerisce l’importanza del sistema linfatico nel drenaggio del FCS durante questo periodo iniziale di sviluppo [13,14].
Nella senescenza, i VA si occludono, degenerano e la membrana aracnoidea s’ispessisce. Ciò si verifica insieme a cambiamenti degenerativi nel plesso coroideo. Tali cambiamenti nel sistema del FCS provocano un ristagno del FCS e svolgono probabilmente un ruolo importante nella demenza delle persone anziane [15,16].
Caratteristiche morfologiche dei villi aracnoidei cranici
La correlazione tra il nuovo concetto di un percorso aperto tra il FCS ed il sangue del seno venoso e la conformazione morfologica presentata da Welch e colleghi, utilizzando metodi con microscopi ottici, non fu universalmente accettata [5,10]. Questo nuovo concetto di drenaggio del FCS con percorso aperto è stato valutato usando sia microscopi elettronici a trasmissione (TEM) che microscopi elettronici a scansione (SEM) in una serie di studi sia sui primati che sugli animali inferiori. Tali studi hanno proposto opinioni contrastanti sulla natura della correlazione tra sangue venoso e FCS. Gli studi precedenti che utilizzavano microscopia ottica ed avanzata sembravano sostenere la tesi secondo la quale non c’era alcuna discontinuità dell’endotelio del seno e pertanto qualsiasi trasferimento di FCS nel sangue attraverso i VA sarebbe avvenuto necessariamente attraverso le cellule endoteliali intatte, anche se studi ripetuti hanno mostrato alcune schisi interendoteliali che hanno permesso la penetrazione di perossidasi di rafano [3,17]. Nelle pecore e nei cani, Gomez et al, hanno osservato schisi intercellulari endoteliali attraverso l’aumento delle pressioni dei liquidi nel seno e nel FCS [18]. La pinocitosi è stata anch’essa osservata e dipendeva in modo simile dalla pressione. Tripathi [19] dimostrò che quando il tessuto era fissato a pressione differenziale bassa o negativa tra lo spazio subaracnoideo ed il seno venoso cranico c’era continuità della superficie endoteliale sopra l’interfaccia aracnoidea con molti microvilli. Quando si è verificato il fissaggio del tessuto a pressione normale ed aumentata, sono stati osservati grandi vacuoli intracellulari, che hanno mostrato una connessione aperta in evoluzione tra il FCS ed il sangue venoso. Tripathi (1968) descrisse inizialmente tale concetto della vacuolizzazione delle cellule endoteliali nel canale di Schlemm, ma lo applicò in seguito al sistema di deflusso dei VA (figura 4) [3,20]. L’apertura dei vacuoli come un grande poro sulla superficie delle cellule endoteliali è dimostrata nella figura 5. Nella figura 6, le microfotografie elettroniche dei VA provano il collegamento di questi vacuoli con lo spazio subaracnoideo e le lacune durali. Tali osservazioni istologiche sono state confermate successivamente da Levine et al [21]. Si pensava che i vacuoli intracellulari fossero la via principale per il drenaggio di massa del FCS e del particolato. I pori aracnoidei, tuttavia, non sono stati visti sull’inversione del gradiente di pressione usato nel fissaggio del tessuto.
Sulla base dei loro studi istologici su materiale umano post mortem e della rispettiva revisione della letteratura, Upton e Weller [22] credevano che l’anatomia dei VA di animali e primati fosse leggermente diversa e che si dovesse pertanto essere cauti nell’estrapolazione tra le specie. Essi hanno notato soprattutto il ruolo significativo della pinocitosi in animali inferiori rispetto al ruolo dei vacuoli transcellulari nella scimmia e nell’uomo. In generale, la letteratura ha sostenuto il concetto secondo il quale il drenaggio del FCS attraverso le granulazioni aracnoidee avviene per pinocitosi endoteliale e vacuolizzazione, nonché attraverso cisterne extracellulari [23]. Negli studi più recenti, ottenuti mediante l’uso di un modello di membrana della granulazione aracnoidea in vitro o ex vivo, derivato da campioni umani post mortem, Grzybowski et al [24] e Glimcher et al [25] hanno dimostrato la base morfologica dei percorsi del FCS in questi trapianti biologicamente vitali conformi agli studi precedenti di cui sopra.
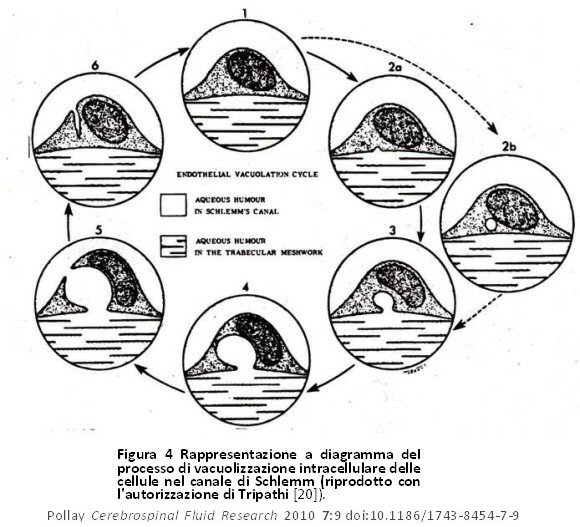 Figura 4 Rappresentazione a diagramma del processo di vacuolizzazione intracellulare delle cellule nel canale di Schlemm (riprodotto con l’autorizzazione di Tripathi [20]). Figura 4 Rappresentazione a diagramma del processo di vacuolizzazione intracellulare delle cellule nel canale di Schlemm (riprodotto con l’autorizzazione di Tripathi [20]). |
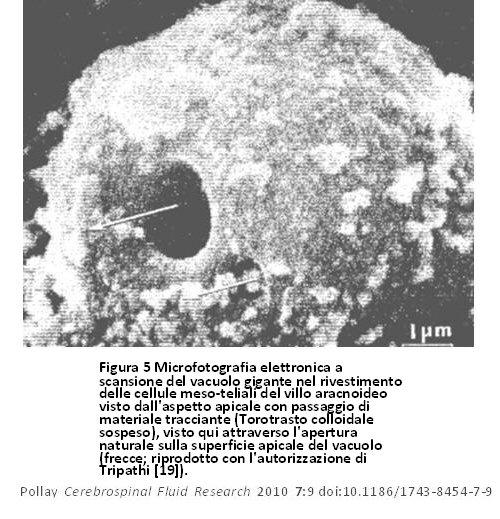 Figura 5 Microfotografia elettronica a scansione del vacuolo gigante nel rivestimento delle cellule meso-teliali del villo aracnoideo visto dall’aspetto apicale con passaggio di materiale tracciante (Torotrasto colloidale sospeso), visto qui attraverso l’apertura naturale sulla superficie apicale del vacuolo (frecce; riprodotto con l’autorizzazione di Tripathi [19]). Figura 5 Microfotografia elettronica a scansione del vacuolo gigante nel rivestimento delle cellule meso-teliali del villo aracnoideo visto dall’aspetto apicale con passaggio di materiale tracciante (Torotrasto colloidale sospeso), visto qui attraverso l’apertura naturale sulla superficie apicale del vacuolo (frecce; riprodotto con l’autorizzazione di Tripathi [19]). |
 Figura 6 Microfotografia elettronica delle cellule mesoteliali di rivestimento del villo aracnoideo che mostra un vacuolo vuoto gigante (V). Il vacuolo a destra presenta sia un’apertura basale che un’apertura apicale, costituendo così un canale vacuolare transcellulare (freccia lunga). Il vacuolo a sinistra presenta solo un’apertura basale (frecce corte). SAS: spazio subaracnoideo; L: lacune della dura (riprodotta con l’autorizzazione di Tripathi [19]). Figura 6 Microfotografia elettronica delle cellule mesoteliali di rivestimento del villo aracnoideo che mostra un vacuolo vuoto gigante (V). Il vacuolo a destra presenta sia un’apertura basale che un’apertura apicale, costituendo così un canale vacuolare transcellulare (freccia lunga). Il vacuolo a sinistra presenta solo un’apertura basale (frecce corte). SAS: spazio subaracnoideo; L: lacune della dura (riprodotta con l’autorizzazione di Tripathi [19]). |
Studi dinamici del deflusso del FCS attraverso i villi aracnoidei
Come accennato in precedenza, Welch e colleghi hanno dimostrato, in un sistema di perfusione della granulazione aracnoidea isolata, l’esistenza di un flusso unidirezionale dalle meningi al seno e praticamente nessun flusso nella direzione opposta (Figure 2 e 3). È stato inoltre scoperto che le particelle con dimensioni fino a 7,5 μm sono passate attraverso l’interfaccia FCS-sangue [9]. In una serie di studi approfonditi effettuati sui VA e la membrana aracnoidea raccolti, Grzybowski e soci [24,25] hanno misurato la conduttività idraulica cellulare a pressioni fisiologiche normali (Tabella 1). Gli esperimenti hanno dimostrato che sia gli studi in vitro, sia quelli successivi che utilizzavano un preparato tissutale ex vivo presentavano un flusso direzionale simile a quello riscontrato nel modello sperimentale di Welch e Friedman [5,24]. Inoltre, hanno scoperto che i loro campioni di occlusione della membrana erano biologicamente vitali e stabili durante lo studio di perfusione. Usando un modello ex vivo, hanno altresì dimostrato che il movimento delle particelle (da 0,1 a 2,0 μm) attraversava la membrana nella direzione fisiologica. L’effettiva legittimazione di questa metodologia sperimentale dovrebbe consentire una comprensione più approfondita del trasferimento di numerose sostanze biologicamente importanti dal FCS alla circolazione sistemica.
Tabella 1: Riassunto delle granulazioni aracnoidee perfuse in vitro nel fisiologico (direzione da B ad A) e non-fisiologico (direzione da A a B).
È stato dimostrato che il differenziale di pressione necessario per drenare il FCS dallo spazio subaracnoideo nel seno sagittale superiore tramite i VA varia tra 3 e 5 mm Hg. Negli esseri umani, utilizzando l’infusione di FCS artificiale, la pressione nel seno sagittale (SSVP) rimane costante su un’ampia gamma di pressioni del FCS artificialmente aumentate (CSFP) [3]. La gamma rientrava ampiamente in quanto clinicamente dimostrato. Shulman et al, in un modello canino, hanno dimostrato che in condizioni normali la CSFP, la SSVP e la pressione venosa torculare (TVP) medie erano rispettivamente 147, 90,3 e 46 mm H2O [26]. La SSVP è quindi circa il 60% della CSFP nell’animale normale. Nel cane idrocefalico, il rapporto tra la CSFP e la SSSP è di 0,98, mentre nell’animale normale è di 0,61. In queste condizioni, il rapporto TVP/CSFP cambia lievemente, il che è probabilmente dovuto alla protezione offerta dal rivestimento del torculare nell’osso del cane. Sono stati effettuati numerosi studi relativi alla pressione del FCS ed all’assorbimento del FCS nell’uomo e negli animali in condizioni normali e con idrocefalo. Sia gli studi di Mann et al nei ratti che quelli di McComb et al nei conigli, utilizzando rispettivamente inulina radiomarcata ed albumina sierica, hanno dimostrato la mancanza di movimento del FCS negli emisferi cerebrali o nel seno sagittale superiore a pressioni fisiologiche del FCS [27,28]. Si dovrebbe considerare che negli animali utilizzati in questi studi, i VA erano generalmente assenti o scarsi nei siti del seno sagittale superiore, ma sono stati osservati principalmente alla base del cranio. Tali risultati saranno discussi in relazione al sistema di drenaggio del nervo olfattivo dopo aver presentato l’anatomia e la fisiologia del sistema di deflusso linfatico.
Conseguenze funzionali dei cambiamenti morfologici nei villi aracnoidei
Anche se è stato dimostrato che l’idrocefalo può essere una conseguenza d’infezioni delle meningi, emorragia e infusione subaracnoidea di FGF-2 (fattore di crescita dei fibroblasti) nel FCS, non è chiaro se le alterazioni nella circolazione e nel drenaggio del FCS siano dovute a cambiamenti patologici nelle vie subaracnoidee o a talune modifiche del duplice sistema di deflusso (VA e linfatico). Negli studi post mortem sulla risposta dei VA all’emorragia subaracnoidea, si è constatato che la proliferazione osservata delle cellule aracnoidee innescata da una reazione infiammatoria o prodotti di coagulazione del sangue può provocare un’ostruzione del flusso di FCS attraverso i VA all’interno dei seni venosi del cranio [29]. Non è possibile escludere l’effetto sulle vie subaracnoidee o sul drenaggio nel sistema linfatico olfattivo in seguito ad un’emorragia subaracnoidea. Tale difficoltà è osservata anche nella valutazione del ruolo specifico dei cambiamenti patologici nello spazio subaracnoideo, nei VA e nei vasi linfatici nello sviluppo dell’idrocefalo causato da infezione alle meningi. Tra le conseguenze funzionali dell’assenza totale, o quasi totale, dei VA negli esseri umani si è riscontrato lo sviluppo dell’idrocefalo [30].In questi casi comprovati, lo sviluppo dei percorsi utilizzati dal sistema linfatico e dai VA spinali non era stato valutato, pertanto anch’essi possono aver giocato un ruolo nella sua determinazione. Lo stesso problema esiste quando l’assorbimento del FCS è influenzato da un aumento marcato della viscosità del liquido circolante (es. alto contenuto proteico). In altre parole, l’assorbimento alterato potrebbe essere a livello delle vie dei VA o di quelle subaracnoidee e linfatico-perineurali.
Ci sono inoltre cambiamenti morfologici e funzionali nei VA che influenzano l’assorbimento del FCS con conseguenti cambiamenti variabili nella pressione intracranica (ICP).Negli studi su animali immaturi, si è notato che la carenza di vitamina A può portare all’ispessimento ed all’infiltrazione della dura madre intorno ai VA con mucopolisaccaridi. Ciò sembra essere associato all’assorbimento diminuito del FCS ed a quello aumentato dell’ICP, a causa dello sviluppo dell’idrocefalo [31]. È stato dimostrato che l’effetto negativo dell’ipovitaminosi A in studi di perfusione ventricolo-cisternale nel modello bovino è dovuto direttamente alla diminuzione dell’assorbimento dei VA [32]. È stato documentato nei bambini che l’ipovitaminosi secondaria può portare ad un aumento dell’ICP e ad un rigonfiamento della fontanella [33]. Livelli tossici di vitamina A possono inoltre influire sulla morfologia e la funzionalità dei VA, del plesso coroideo e degli epiteli ependimali. Negli animali colpiti, le dimensioni dei VA si sono ridotte rispetto ai controlli e il cappuccio aracnoideo era più sottile e più piccolo. Si è suggerito che questi cambiamenti fossero associati ad un aumento del deflusso del FCS con alterazione della morfologia funzionale del plesso coroideo con una conseguente bassa ICP [34]. In generale, i livelli tossici di retinolo nel FCS possono determinare una sindrome da ipertensione intracranica idiopatica [35].
I villi aracnoidei spinali
I primi studi sui VA spinali hanno dimostrato la presenza di cluster e proiezioni aracnoidei verso e attraverso la dura madre nella regione dei gangli della radice dorsale, ma non li hanno pienamente correlati ai vasi venosi della regione o alla possibilità che queste strutture abbiano rappresentato un luogo di deflusso del FCS [3]. Elman pensò che queste strutture fossero simili ai villi cranici e notò il rapporto con le strutture venose della zona [7]. Wislocki ritenne che questi seni venosi spinali fossero equivalenti ai seni venosi cranici e notò la relazione speciale con i villi aracnoidei [8]. Brierley e Fields, utilizzando un tracciante a base d’inchiostro di china iniettato nel FCS, dimostrarono che il tracciante aveva raggiunto lo spazio epidurale spinale e i vasi linfatici della regione [36]. Nel 1961, Welch e Pollay rivisitarono, nei cercopitechi gialloverdi, il rapporto anatomico dei VA della radice dorsale e le vene della regione [37]. Nella figura 7, viene mostrata la natura molteplice di tale rapporto. Da tale studio sui cercopitechi gialloverdi è risultato chiaro che i VA penetravano più comunemente nello spazio epidurale, ma si è inoltre osservato che venivano penetrate le vene con pareti sottili intorno ai gangli della radice dorsale. Queste scoperte e gli studi precedenti che utilizzavano il trasferimento del tracciante dallo spazio subaracnoideo spinale alle strutture vascolari adiacenti supportano la base anatomica del sistema di deflusso dei VA spinali. L’importanza dei vasi linfatici nel deflusso cranico e spinale non è stata pienamente valutata a quel tempo. Nella scimmia, i VA spinali trovati in correlazione con le strutture venose sono stati osservati sono nel 16% delle radici studiate. Gomez et al hanno riscontrato un’incidenza leggermente maggiore di VA spinali nei cani e nelle pecore [38]. Kido et al scoprirono nei cadaveri umani che la concentrazione di VA spinali era più comune nella regione toracica [39]. Tubbs et al rivisitarono il rapporto tra i VA spinali e le strutture venose adiacenti nei cadaveri umani adulti [40]. Non trovarono VA su tutte le radici, ma la maggior concentrazione fu riscontrata nella regione lombare, sempre in stretta correlazione con i vasi venosi della regione. C’era anche un rapporto diretto tra la dimensione della vena radicolare adiacente e la presenza ed il numero dei VA.
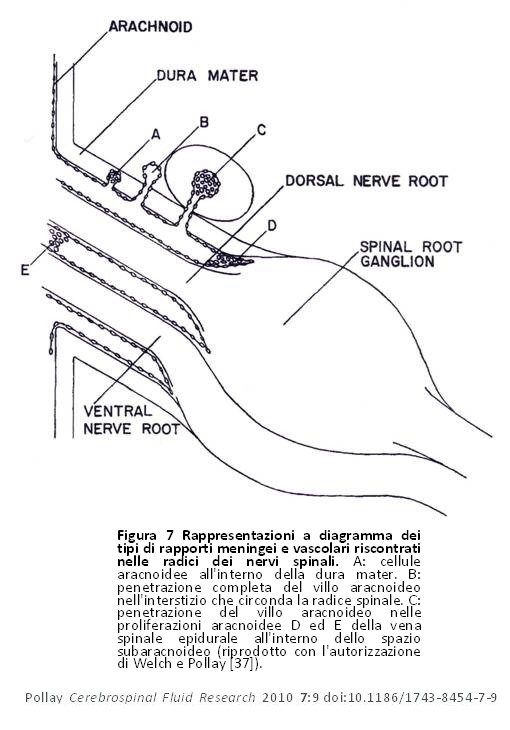 Figura 7 Rappresentazioni a diagramma dei tipi di rapporti meningei e vascolari riscontrati nelle radici dei nervi spinali. A: cellule aracnoidee all’interno della dura mater. B: penetrazione completa del villo aracnoideo nell’interstizio che circonda la radice spinale. C: penetrazione del villo aracnoideo nelle proliferazioni aracnoidee D ed E della vena spinale epidurale all’interno dello spazio subaracnoideo (riprodotto con l’autorizzazione di Welch e Pollay [37]). Figura 7 Rappresentazioni a diagramma dei tipi di rapporti meningei e vascolari riscontrati nelle radici dei nervi spinali. A: cellule aracnoidee all’interno della dura mater. B: penetrazione completa del villo aracnoideo nell’interstizio che circonda la radice spinale. C: penetrazione del villo aracnoideo nelle proliferazioni aracnoidee D ed E della vena spinale epidurale all’interno dello spazio subaracnoideo (riprodotto con l’autorizzazione di Welch e Pollay [37]). |
Marmarou et al [41] lavorando sui gatti, separarono il compartimento cranico da quello spinale con un palloncino gonfiabile e scoprirono che il compartimento spinale rappresentava il 16% dell’assorbimento del FCS e che l’assorbimento spinale sembrava simile all’assorbimento cranico per il fatto che non c’erano differenze distinguibili in base alle dinamiche di pressione. Nelle pecore, è stato dimostrato che il rapporto della clearance cranico/spinale dello iodio radiomarcato nel compartimento del FCS variava a seconda del metodo usato nello studio, ovvero l’iniezione di bolo o l’infusione cranica e spinale [42]. Espresso come percentuale del drenaggio totale del FCS, il rapporto della clearance cranico/spinale per perfusione, iniezione di bolo e introduzione tramite serbatoio in compartimenti sia cranici che spinali, è stato stimato rispettivamente a 75:25, 88:12 e 75:25. Si è concluso che circa il 25% del drenaggio del FCS avviene attraverso i VA spinali. Negli esseri umani, utilizzando la cisternografia con puntura lombare e con radionuclide, Edsbagge et al [43] hanno calcolato la formazione ed il drenaggio di FCS misurando l’attività spinale del radionuclide in esseri umani giovani ed in salute. Il tasso di diminuzione dell’attività del tracciante è stato circa del 20% nella prima ora ed è migliorato con l’attività fisica nei soggetti umani testati. La produzione media di FCS era di circa 0,35 ml/min. Sulla base della riduzione dell’attività del nuclide nel compartimento spinale, l’assorbimento spinale si attestava da 0,11 a 0,23 ml/min, con un afflusso netto di liquor ventricolare nello spazio subaracnoideo di 0,45 e 0,48 ml/min [43]. Tali valori si sono rivelati un po’ più alti rispetto a quanto precedentemente indicato. La questione irrisolta rimaneva il fatto che il drenaggio attraverso i VA avvenisse all’interno dei vasi linfatici che si trovano intorno alle radici spinali o nelle vene in cui alcuni VA penetrano. La maggior parte degli studi con traccianti sostiene che il collegamento linfatico sia più importante in tale configurazione [36,42].
La storia antica dello studio sul drenaggio linfatico del fluido cerebrospinale
Sebbene Schwab (1869) avesse osservato che le sostanze iniettate nello spazio subaracnoideo, in un secondo momento, potessero trovarsi nei linfonodi cervicali, furono Key e Retzius a dimostrare il percorso utilizzato dal tracciante per raggiungere il sistema linfatico cervicale [4].
Per i 30 anni successivi, numerosi autori hanno dimostrato in animali viventi, utilizzando molteplici traccianti colorati o radiomarcati, che c’era un percorso lungo i nervi olfattivi ed ottici, che alla fine portava all’accumulo di traccianti nei linfonodi cervicali [3,44].
Utilizzando la visualizzazione a raggi x, l’uscita di torotrasto e brominal è stata dapprima mostrata da Mortensen e Sullivan (1933) e, in seguito, da Faber (1937), rispettivamente nel cane e nel coniglio vivente, indicando che il passaggio avveniva attraverso i nervi olfattivi nel sistema linfatico cervicale solitamente entro un periodo di cinque ora dall’iniezione subaracnoidea [45,46]. Iniettando dell’Au198 nel tessuto sub-mucosale del coniglio, si scoprì che penetrava attraverso la lamina cribiforme all’interno delle cisterne basali e nel lobo frontale [47]. Jackson et al [48] presentarono uno schema anatomico di un modello di manicotto perineurale aperto e chiuso per il drenaggio del FCS nel sistema linfatico cervicale tramite lo spazio perineurale olfattivo (figura 8). Allo stesso tempo, è stato dimostrato sperimentalmente nel coniglio e nel gatto, che il sistema linfatico è in grado di rimuovere sia le proteine che i globuli rossi dal FCS [49,50]. Circa 25 anni più tardi, una serie di studi effettuati sui gatti e sui conigli ha confermato la prima ricerca riguardo all’importanza dei vasi linfatici nel drenaggio del FCS. Cserr et al hanno osservato l’importanza del percorso linfatico nel coniglio, nella pecora e nel gatto utilizzando un tracciante radiomarcato all’albumina iniettato nel FCS o direttamente nel cervello [51]. Hanno scoperto che tra il 14 e il 47% del tracciante iniettato nel cervello passa attraverso il sistema linfatico. Hanno inoltre suggerito che il sistema di deflusso linfatico del FCS potrebbe rappresentare il braccio afferente della risposta immunitaria agli antigeni.
Questi autori lasciarono intendere che c’era una comunicazione continua ed altamente regolata tra il cervello e il sistema immunitario [52]. In un modello di ratto, Weller et al hanno inoltre dimostrato l’importanza del sistema di drenaggio linfatico nel drenaggio di fluido dallo spazio interstiziale della corteccia [53]. Sembra che il drenaggio del liquido interstiziale cerebrale nel FCS avvenga tramite i canali perivascolari e che successivamente entri nel sistema linfatico attraverso la lamina cribiforme (figura 9). Ciò supporta la tesi secondo la quale questo percorso svolge un ruolo primario nel ricambio del fluido interstiziale cerebrale. Ci furono inoltre diversi studi che propongono siti potenziali di drenaggio lungo molti degli altri nervi cranici [51,52,54–56]. Essi sono mostrati schematicamente nella figura 10. Bradbury e Cole osservarono una concentrazione significativa di I125 -albumina nei linfonodi cervicali profondi in seguito ad una sola iniezione nel ventricolo laterale, una quantità abbastanza considerevole sia nel coniglio (14,4%) che nel gatto (12,9%) [56]. Il passaggio del tracciante nell’orbita si è anch’esso verificato, ma era quantitativamente poco significativo o nullo e non è stato trovato nei vasi linfatici cervicali dopo 6 ore. In un articolo successivo, Bradbury et al [57] presentarono uno schema del percorso ipotetico del drenaggio di FCS di un tracciante della proteina iniettato nel cervello, per poi entrare nel FCS subaracnoideo mediante gli spazi perivascolari (figura 11). Da questi studi precedenti risulta che il drenaggio del liquido interstiziale del cervello avvenga principalmente, se non esclusivamente, attraverso il sistema di deflusso linfatico nasale e della relativa importanza non solo nella modulazione del volume di liquido cerebrale, ma anche nel fornire una connessione altamente regolata tra il cervello ed il sistema immunitario [51,52,57]. Il ricambio del fluido interstiziale cerebrale rappresenta circa il 10% del ricambio totale del compartimento del FCS [3]. Il tasso di trasferimento del FCS dal compartimento del FCS al sistema di deflusso linfatico è negativamente influenzato anche da una posizione a testa alta dell’animale, dall’ostruzione della zona cribiforme e da traccianti con peso molecolare più elevato [58].
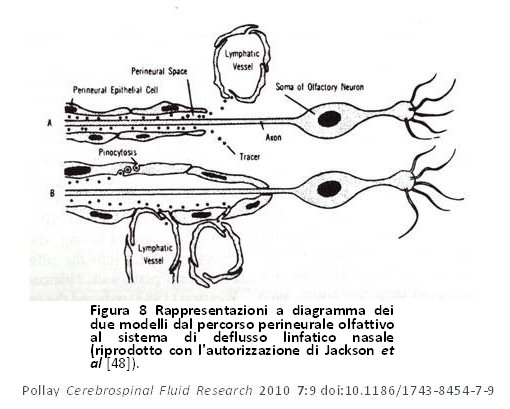 Figura 8 Rappresentazioni a diagramma dei due modelli dal percorso perineurale olfattivo al sistema di deflusso linfatico nasale (riprodotto con l’autorizzazione di Jackson et al [48]). Figura 8 Rappresentazioni a diagramma dei due modelli dal percorso perineurale olfattivo al sistema di deflusso linfatico nasale (riprodotto con l’autorizzazione di Jackson et al [48]). |
 Figura 9 Diagramma schematico che illustra un modello di ricambio del fluido interstiziale nel cervello basato sulla secrezione di ISF cerebrale dalla barriera emato-encefalica (frecce aperte) e flusso di massa di ISF dal cervello al FCS tramite spazi perivascolari (frecce curve). Il FCS è secreto dal plesso coroideo (frecce aperte) e drena con l’ISF dallo spazio subaracnoideo nel sangue venoso e nella linfa (riprodotto con l’autorizzazione di Cserr et al [51]). Figura 9 Diagramma schematico che illustra un modello di ricambio del fluido interstiziale nel cervello basato sulla secrezione di ISF cerebrale dalla barriera emato-encefalica (frecce aperte) e flusso di massa di ISF dal cervello al FCS tramite spazi perivascolari (frecce curve). Il FCS è secreto dal plesso coroideo (frecce aperte) e drena con l’ISF dallo spazio subaracnoideo nel sangue venoso e nella linfa (riprodotto con l’autorizzazione di Cserr et al [51]). |
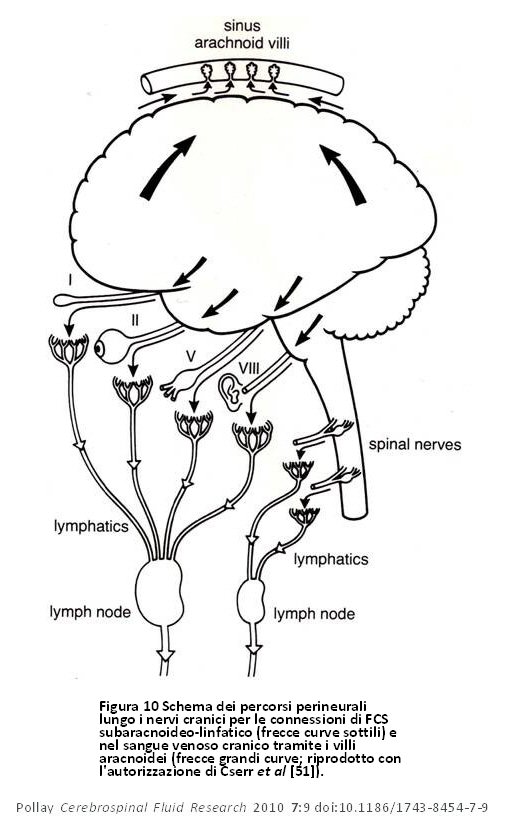 Figura 10 Schema dei percorsi perineurali lungo i nervi cranici per le connessioni di FCS subaracnoideo-linfatico (frecce curve sottili) e nel sangue venoso cranico tramite i villi aracnoidei (frecce grandi curve; riprodotto con l’autorizzazione di Cserr et al [51]). Figura 10 Schema dei percorsi perineurali lungo i nervi cranici per le connessioni di FCS subaracnoideo-linfatico (frecce curve sottili) e nel sangue venoso cranico tramite i villi aracnoidei (frecce grandi curve; riprodotto con l’autorizzazione di Cserr et al [51]). |
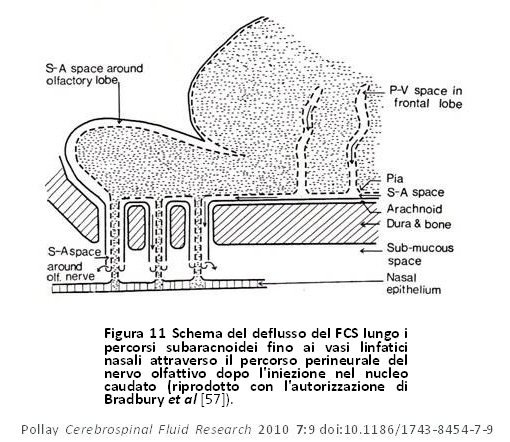 Figura 11 Schema del deflusso del FCS lungo i percorsi subaracnoidei fino ai vasi linfatici nasali attraverso il percorso perineurale del nervo olfattivo dopo l’iniezione nel nucleo caudato (riprodotto con l’autorizzazione di Bradbury et al [57]). Figura 11 Schema del deflusso del FCS lungo i percorsi subaracnoidei fino ai vasi linfatici nasali attraverso il percorso perineurale del nervo olfattivo dopo l’iniezione nel nucleo caudato (riprodotto con l’autorizzazione di Bradbury et al [57]). |
Quasi un decennio sarebbe passato prima che ulteriori elementi di prova quantitativa fossero disponibili a conferma dell’importanza del sistema di deflusso linfatico del FCS nel mantenere l’ambiente cerebrale e la sua rilevanza nel deflusso globale del FCS rispetto al ruolo dei VA [44].
Morfologia funzionale del sistema di drenaggio linfatico del FCS
Considerazioni anatomiche
I primi studi qualitativi hanno suggerito che la base anatomica del passaggio dei traccianti consistesse degli spazi perineurali di alcuni nervi cranici, che all’uscita del cranio permettevano la circolazione del FCS, proveniente dallo spazio subaracnoideo basale, l’entrata nel tessuto interstiziale o direttamente nel sistema linfatico cervicale. Quest’opinione era generalmente limitata al drenaggio del FCS nei vasi linfatici nasali tramite la guaina perineurale del nervo olfattivo dopo la penetrazione nella lamina cribiforme, sebbene siano state trovate informazioni incomplete sui nervi cranici oculari o uditivi [54-56]. Studi ultrastrutturali del nervo olfattivo del coniglio, dopo l’infusione di perossidasi di rafano (HRP), hanno rivelato che l’HRP si spostava lungo lo spazio perineurale e confluiva nei vasi linfatici, nelle ghiandole mucose, negli spazi intercellulari dell’epitelio nasale e nelle venule [59]. Nell’esame post mortem sugli esseri umani a seguito di emorragia subaracnoidea, questi spazi perineurali contenevano globuli rossi sia distalmente che prossimalmente ai nervi olfattivi. Sono stati trovati pigmenti di ferro anche nei linfonodi cervicali [60]. Kida et al hanno trovato particelle di carbonio lungo lo spazio subaracnoideo fino alla cavità nasale, all’orbita ed alla coclea, ma solo la via nasale ha mostrato una comunicazione diretta del FCS cranico con il sistema linfatico [61]. Utilizzando l’HRP come marker, è stata dimostrata una connessione tra gli spazi subaracnoidei ed i vasi linfatici della scala vestibuli[55]. L’anatomia del percorso lungo il nervo ottico è meno chiara, ma utilizzando l’infusione di materiale di contrasto è stato dimostrato che non c’è dispersione all’estremità distale della guaina del nervo ottico (ONS). All’estremità (distale) della parte terminale di questa guaina, lo strato di cellule neuroendoteliale è abbastanza sottile e ci sono diversi filamenti intercellulari ed aperture simili a pori [62].
In seguito all’iniezione di HRP nella teca spinale, Gomez et al, hanno osservato l’HRP nello spazio subaracnoideo intorno al nervo ottico dove è penetrato all’interno del nervo, occupando gli spazi extracellulari tra gli elementi nervosi, fino alla zona della lamina cribosa. Dopo aver attraversato la sclera perineurale, si è diffuso nella coroide ed è stato reperito nei capillari della coroide, nelle venule e nelle vene attraversando gli spazi interendoteliali [62]. Nel cane, l’infusione di colorante blu di Evans nel FCS della cisterna magna rivela il drenaggio nell’orbita con conseguente proptosi [54]. Ciò si sarebbe presumibilmente verificato attraverso l’umore acqueo-canale di Schlemn e il dotto nasolacrimale [63].
La prova più convincente che definisce la base anatomica del sistema di deflusso linfatico nasale del FCS è stata dimostrata da Johnston et al che hanno seguito il percorso di deflusso di microfill colorato iniettato nel FCS [64]. Questi preparati anatomici sono stati testati su suini, conigli, ratti, topi, scimmie e sette giorni dopo la morte negli esseri umani. Il sistema di drenaggio linfatico del FCS è stato identificato dopo l’iniezione di microfill giallo nel compartimento del FCS. Negli animali studiati, il microfill ha riempito lo spazio subaracnoideo e si è inserito nella rete dei vasi linfatici nella sottomucosa nasale. In genere, negli animali inferiori, il microfill non è entrato nell’interstizio della mucosa nasale, ma si è inserito nei vasi linfatici direttamente dallo spazio subaracnoideo perineurale. L’effettiva configurazione di quest’interfaccia non era chiara, ma la disposizione più probabile è un collegamento diretto con i vasi linfatici, poiché nessun microfill è stato trovato nell’interstizio [64]. Negli esseri umani, ci sono state alcune extravasazioni nello spazio interstiziale della mucosa nasale che avrebbero potuto essere principalmente dovute al deterioramento dei campioni post mortem. Questo studio ha stabilito con fermezza la base anatomica del sistema di deflusso linfatico, che era stato precedentemente dimostrato dai primi studi semi-quantitativi [3,48,51] e dai successivi studi volumetrici [44]. L’effettiva configurazione microscopica dell’interfaccia tra lo spazio perineurale e la parete o il lume dei vasi linfatici rimane sconosciuta. Con livelli di pressione fisiologica, il microfill iniettato non è riuscito a dimostrare l’entrata nel sistema venoso cranico attraverso i VA, anche se questo non è accaduto quando la pressione del FCS veniva aumentata.
Analisi volumetrica del sistema di drenaggio linfatico del FCS
Kida et al [65] hanno confermato nei loro studi sui ratti che, dopo l’iniezione di inchiostro di china nella cisterna magna, questo tracciante è principalmente rimosso dal sistema linfatico e in misura minore dai VA. Questa conclusione è stata supportata da prove anatomiche e non fisiologiche. Tale opinione costante era generalmente supportata negli esseri umani da esami post mortem in casi fatali di emorragia subaracnoidea in cui i globuli rossi sono stati trovati sia nel sistema linfatico nasale che nei VA. Bradbury et al scoprirono che la percentuale della dose di tracciante iniettata nel FCS trovato nel sistema linfatico cervicale profondo rappresentava quasi il 30% del FCS prodotto nel coniglio ed il 10-15% nel gatto [3,57]. Johnston e i suoi colleghi hanno indagato sulle capacità funzionali del sistema linfatico in una serie di pubblicazioni distinte basate su esperimenti volti a fornire una valutazione quantitativa dei vari componenti del sistema di drenaggio totale del FCS nelle pecore e nei ratti. Gli studi volumetrici sono stati effettuati principalmente in pecore coscienti in cui un tipo di iodio radiomarcato era utilizzato per misurare il deflusso di FCS tramite le vie nasali linfatiche e il sistema di deflusso cranico e spinale [66,67]. Il secondo tipo di iodio radiomarcato era utilizzato per correggere la filtrazione del marker di FCS raggiungendo il sistema linfatico dal sangue per tornare nel plasma da siti di drenaggio non-linfatico. È stato ipotizzato nelle equazioni derivate che i VA ed il percorso linfatico fossero gli unici siti di assorbimento del sistema del FCS. Era ragionevole supporre che nessuna perdita significativa di tracciante si fosse verificata attraverso la barriera emato-encefalica o il plesso coroideo. Il modello matematico era basato su un modello a tre compartimenti semplice ma ragionevole (FCS, plasma e linfa) e prevedeva variazioni tra animali a concentrazioni di isotopi in vari compartimenti e costanza o affidabilità delle disposizioni anatomiche. Negli animali coscienti, utilizzati in questi esperimenti, si presumeva che le pressioni all’interno dei compartimenti del FCS e il tasso di produzione di liquido e il flusso sanguigno fosse rimasto entro i limiti fisiologici. Hanno confermato l’opinione convenzionale secondo la quale una porzione del FCS entra nei linfonodi cervicali superficiali e profondi. Il reperimento del tracciante nei nodi lombari e intercostali ha indicato la presenza di un certo drenaggio spinale di FCS [66,67]. Il recupero dell’albumina sierica umana radiomarcata (HSA) sia nei sistemi di drenaggio linfatico che nei VA è stato effettuato mediante il campionamento dei vasi linfatici cervicali cannulati e del dotto toracico, nonché del sangue venoso. A 6 ore dall’iniezione nel ventricolo cerebrale, l’8,2% ed il 12,5% della dose iniettata è stato recuperato rispettivamente dal deflusso linfatico e dai VA. Dopo 22 ore, il recupero era del 25,1% e del 20,8%. Questi risultati hanno dimostrato che la clearance del tracciante era quasi equamente distribuita tra i sistemi di drenaggio linfatico e i VA nel rispettivo modello animale [66].Non c’era alcuna differenza significativa nella quantità recuperata nei due sistemi di deflusso quando il tracciante è stato iniettato nel FCS lombare. In uno studio successivo, il tasso totale di assorbimento del FCS (percorsi linfatico e dei VA) poteva essere stimato a 3,48 ml/h [67].
Boulton et al studiarono l’effetto della pressione sulla clearance del tracciante dal sistema ventricolare. Usarono una disposizione sperimentale simile in cui la raccolta del tracciante radiomarcato dai sistemi linfatico e venoso è stata valutata durante una perfusione di 3 ore a bassa pressione e per un periodo simile per la valutazione del deflusso a vari livelli di pressione [68]. I risultati sono mostrati nella tabella 2. In media, un aumento dell’ICP di 10 cm H2O ha aumentato la clearance del FCS dei VA e del sistema linfatico rispettivamente di 2,7 e 3,9 volte.
Tabella 2: effetto della pressione sul deflusso del fluido cerebrospinale dal sistema ventricolare
L’effetto dell’ostruzione dei vasi linfatici cervicali e dello spazio subaracnoideo spinale sul drenaggio del FCS
Quando i vasi linfatici cervicali e i nodi cervicali erano legati, sono state riportate conseguenze intracraniche significative come l’aumento dell’ICP, dell’EEG e modifiche comportamentali negli animali testati [44]. Tali studi precedenti hanno suggerito l’importanza del sistema linfatico nel mantenimento di una normale omeostasi del liquido intracranico. La clearance dei traccianti dal FCS attraverso il sistema linfatico nasale in seguito all’ostruzione della lamina cribiforme nei ratti ha rivelato che la dose recuperata era lo 0,697% prima della legatura linfatica e dello 0,357% dopo la legatura. Si è concluso che circa il 50% del deflusso è avvenuto tramite il sistema linfatico e il resto a causa del drenaggio cranico e dei VA spinali [69].
Per valutare il ruolo del drenaggio linfatico non-nasale del FCS tramite la lamina cribiforme negli ovini, lo spazio subaracnoideo cranico è stato separato dalle vie di deflusso nasali ostruendo la lamina cribiforme e determinando così l’impatto sulla resistenza di deflusso rispetto al drenaggio del FCS. La resistenza di deflusso del FCS è aumentata di circa 2,7 volte e si è notato un innalzamento associato dell’ICP ed un aumento del tempo necessario per la diminuzione della pressione alla base dopo un’iniezione di bolo [70]. Negli ovini neonati, Papaiconomou et al hanno scoperto che la chiusura ermetica della lamina cribiforme spostava la curva di assorbimento del flusso ICP – FCS a sinistra [14]. Quando il compartimento cranico è stato separato dal compartimento spinale, tale effetto su pressione-flusso è stato migliorato. In altre parole, tali pressioni superiori di FCS erano necessarie per gestire l’assorbimento del FCS. Il trasporto del tracciante radiomarcato nel sangue venoso cranico è stato anch’esso significativamente aumentato in queste condizioni. Sembra che tale studio indichi un ruolo importante ma secondario dei VA cranico nel drenaggio del FCS, soprattutto negli ovini neonati con aumento dell’ICP, dopo aver isolato i sistemi di deflusso linfatico e spinale. Tale ruolo secondario potrebbe essere previsto dal momento che c’è una relativa scarsità di strutture dei villi lungo il seno sagittale superiore in questa fascia di età [71,72].
Sviluppo e capacità funzionali del sistema di drenaggio linfatico del FCS
Lo sviluppo di siti di assorbimento del FCS nel ratto e nei suini sono stati studiati in diverse occasioni prima e dopo la nascita da Koh et al [73]. Le connessioni linfatiche-FCS sono state valutate utilizzando microfill colorati ed un complesso della proteina blu di Evan solubile. I suini sono stati scelti perché si verifica una sintesi del FCS prima della nascita, mentre nel ratto la formazione del FCS è marcatamente regolamentata entro un breve periodo di tempo (poche settimane) dalla nascita. Nei suini, la connessione linfatica-FCS non è stata osservata in feti E80-81, ma è stata osservata prima, addirittura a E92. Nei ratti, tali connessioni non sono state riscontrate fino ad una settimana dalla nascita. Sembrerebbe che l’insorgenza della formazione del FCS sia ben correlata con la creazione del sistema di deflusso linfatico in entrambe le specie.
È stato fatto un confronto del drenaggio del FCS tra feti di ovini, ovini neonati ed ovini adulti [14,74]. Una sintesi della resistenza e della conduttanza del deflusso è mostrata nella tabella 3. I valori in tutte le fasce d’età sono abbastanza simili, con la maggiore concentrazione del tracciante radiomarcato HSA riscontrato nei vasi linfatici cervicali. Nei ratti e nei topi, la resistenza del deflusso al drenaggio del FCS aumenta alla nascita e poi diminuisce progressivamente con l’età [75,76]. Si è suggerito che ciò sia dovuto all’ulteriore sviluppo dei VA e correlato all’aumento della formazione del FCS. È stato inoltre osservato che un gradiente di pressione positivo del seno sagittale-FCS non è stato riscontrato, il che suggerisce un ruolo primario dei vasi linfatici in questo primo periodo di tempo. Il tasso del trasporto globale di FCS e la resistenza del deflusso nel periodo neonatale delle pecore si sono dimostrati simili a quelli delle pecore adulte (tabella 3). Tale studio [14] come quello precedente [74], che valutava il feto animale, indica che anche in presenza di meno VA in età precoce, esiste un sistema di deflusso del FCS con una capacità simile a quella dell’adulto. Il cambiamento dell’ICP e della velocità del flusso osservato in età neonatale dopo aver isolato il compartimento cranico del FCS dal deflusso linfatico olfattivo ha suggerito a questi autori che i VA giochino un ruolo limitato nello sviluppo precoce, nonché un ruolo secondario ma più importante nell’animale adulto [44,73,74].
Tabella 3: dinamiche del FCS negli ovini in stato fetale, neonatale ed adulto.
L’impatto dell’invecchiamento sul FCS linfatico nel ratto è stato studiato in 344 ratti Fischer con intervalli di età da 3 a 19 mesi [77]. A 30 minuti dall’iniezione di I-HSA125, la % iniettata/g tessuto dai turbinati nasali era di 6,68 ± 0,42 a 3 mesi, di 4,78 ± 0,67 a 6 mesi, di 2,49 ± 0,31 a 12 mesi e di 2,42 ± 0,72 a 19 mesi. Questo ha rappresentato un calo significativo della capacità di drenaggio del sistema linfatico del FCS. La diminuzione associata alla formazione del FCS a causa dell’età contribuisce anche alla diminuzione del ricambio di FCS riscontrato negli anziani [15].
Idrocefalo e compromissione dell’assorbimento del FCS da parte del sistema linfatico
Nei ratti con idrocefalo a comunicazione indotta mediante caolino, l’HSA radiomarcato è stato iniettato nel ventricolo laterale ed è stato misurato l’arricchimento dei turbinati olfattivi [78]. La presenza del label I 125 nei turbinati era dello 0,99 ± 0,39 (% iniettata/g tessuto) negli animali idrocefalici rispetto a 5,86 ± 0,32 nei controlli. Si è suggerito che la compromissione del percorso subaracnoideo da e per la lamina cribiforme fosse causata o avesse contribuito allo sviluppo dell’idrocefalo. Ciò è stato monitorato in uno studio di Rammling et al che si sono concentrati sulla penetrazione del colorante blu di Evan immediatamente dopo un’iniezione post mortem nella cisterna magna dei ratti H-Tx con idrocefalia congenita [79]. La visualizzazione del colorante nella regione olfattiva dei ratti Sprague-Dawley del controllo e dei ratti da H-Tx affetti e non affetti ha mostrato meno colorante nella regione olfattiva dei ratti H-Tx idrocefalici. Si è concluso, come con l’idrocefalo indotto con caolino, che l’idrocefalo osservato fosse dovuto principalmente all’ostruzione nella porzione olfattiva del sistema di drenaggio linfatico del FCS. Da notare che i VA sono scarsi nei ratti, il che suggerisce il ruolo primario del sistema di deflusso linfatico in tale specie.
Altri siti di assorbimento del FCS
Gli altri siti possibili per l’assorbimento del FCS ??sono la membrana aracnoide, il plesso coroideo e il capillare cerebrale. Usando il modello ex vivo contenente i VA ed il modello adiacente aracnoide. Grzybowski et al, utilizzando un preparato in vitro di VA come membrana di occlusione in un sistema di perfusione, hanno dimostrato delle proprietà di flusso simili a quelle osservate nel preparato di perfusione ex vivo di VA [24,25]. Ciò ha suggerito che tutta la membrana aracnoide fosse in grado di effettuare un drenaggio di massa del FCS [24]. Tuttavia, ciò deve ancora essere dimostrato negli animali intatti. In circostanze normali, è possibile che l’Aquaporin4 trovata all’interfaccia delle cellule gliali-endoteliali influenzi l’assorbimento del fluido interstiziale nel sangue a pressioni fisiologiche, anche se è stato dimostrato in modo convincente che l’assorbimento ed il ricambio del fluido interstiziale cerebrale è gestito adeguatamente dal sistema linfatico nasale [51]. I co-trasportatori di glucosio e di proteine GLUT1, KCC1 e MCT1 si trovano sulle membrane luminali e cerebrali di fronte alle membrane della cellula endoteliale. Anche se la permeabilità osmotica passiva totale all’acqua fornita da questi trasportatori può essere significativa nell’omeostasi del cervello, il loro ruolo nel trasporto netto dell’acqua attraverso la BBB non è ancora ben compreso [80]. La bassa permeabilità della membrana basale e delle giunzioni intercellulari del plesso coroideo, nonché i gradienti idrostatici o osmotici inadeguati rendono improbabile che il movimento di massa del FCS nei capillari del plesso coroideo avvenga in condizioni fisiologiche normali.
Drenaggio del FCS nell’idrocefalo
È stato proposto da Johanson et al che l’aquaporin1 (AQP1) e l’aquaporin4 (AQP4), rispettivamente presso la barriera sangue-FCS e sangue-cervello (cellule endoteliali dei capillari cerebrali), possono essere modificate nell’idrocefalo sperimentale [15]. Una certa compensazione della situazione idrocefalica avverrebbe attraverso la regolazione verso l’alto di AQP4 all’interfaccia gliali-endoteliale che porta al miglioramento del trasporto di liquido interstiziale cerebrale nel sangue, mentre una regolazione verso il basso di AQP1 alla cellula coroidea diminuirebbe la formazione del FCS. Entrambi tenderebbero a migliorare l’accumulo di liquido nell’idrocefalo. Nella comunicazione dell’idrocefalo infiammatorio nei ratti è stato dimostrato che l’aggiornamento dell’AQP4 periventricolare era fortemente correlato alle dimensioni ventricolari. L’AQP4 è stato dapprima localizzato nei piedi terminali degli astrociti ma, successivamente, l’intera membrana degli astrociti è diventata ipertrofica in casi gravi di idrocefalo [81]. Un simile aggiornamento di questa proteina a canale d’acqua è stato riportato nel ratto H-Tx con idrocefalo arrestato spontaneamente, che suggerisce un mezzo di compensazione per assorbimento disturbato del FCS [82]. Non si conosce l’efficacia delle variazioni nell’acquaporina nel miglioramento significativo delle condizioni idrocefaliche, ma suggerisce una risposta protettiva.
È stato dimostrato che l’ostruzione di uno dei siti per il drenaggio del FCS porta all’uso dei siti alternativi rimanenti. Il drenaggio del FCS sia attraverso il sistema di deflusso dei VA (cranico e spinale) che di quello linfatico aumenta considerevolmente con l’aumento della pressione del FCS [44,68]. Ne conseguirebbe quindi che ciò si sarebbe verificato nel sistema intatto dopo che un deflusso del sistema diminuisce con l’aumento della resistenza al deflusso e dell’ICP. Ciò è difficile da dimostrare poiché provare che un processo patologico che colpisce un percorso lascia una via alternativa morfologicamente e/o funzionalmente intatta è veramente difficile. Questo è vero per la maggior parte dei modelli infiammatori (per esempio, caolino, emorragia o infezione) ed alcuni modelli congeniti. Anche se l’emorragia subaracnoidea negli esseri umani può portare a idrocefalo con significative alterazioni fibrotiche nei VA, ci possono essere cambiamenti nella pervietà dello spazio subaracnoideo e prove di accumulo di globuli rossi nei percorsi perineurali olfattivi. È stato notato che l’aumento del TGF-Beta 1 provoca idrocefalo dopo un’emorragia subaracnoidea indotta nei topi tramite iniezione intraventricolare. È emerso che il principale cambiamento patologico consisteva nell’interferenza del flusso subaracnoideo del FCS a causa della maggiore cellularità ed alle alterazioni fibrotiche della leptomeninge. In queste condizioni, è difficile separare la funzione alterata di un sito dagli altri coinvolti nel deflusso del FCS. Negli animali più soggetti ad esperimenti, il lasso di tempo per la maturazione dei vari sistemi di deflusso può anche confondere l’interpretazione della capacità di percorsi alternativi negli animali giovanissimi [14]. La scoperta di un’assenza o scarsità di granulazioni aracnoidee negli esseri umani con idrocefalo suggerisce l’importanza di queste strutture nel drenaggio totale del FCS, ma l’esame post mortem in questi soggetti ha fornito una valutazione incompleta del sistema linfatico, dello spazio subaracnoideo globale e dei VA spinali [30]. In seguito a chirurgia sulla base cranica, l’8% dei pazienti sviluppa idrocefalo nel periodo post-operatorio [83]. In questo gruppo di pazienti, un numero ancora maggiore ha sviluppato perdite di FCS che hanno richiesto uno shunt. Anche in questo caso è difficile isolare il sito responsabile della lesione. L’osservazione del fatto che l’idrocefalo non sia abitualmente riscontrato dopo la rimozione di tumori della fossa anteriore o in seguito alla riparazione delle perdite del FCS dovute a interruzioni di radichette olfattive sovrastanti la zona cribiforme può sostenere l’importanza dei VA cranici nel drenaggio del FCS nei primati ed un ruolo meno importante nel sistema di drenaggio linfatico in tali specie. In caso di scarso sviluppo dei bulbi olfattivi, è stato ipotizzato che la perdita potenziale di neuroni olfattivi potrebbe portare alla perdita del condotto ai vasi linfatici cervicali, conducendo pertanto all’idrocefalo osservato [44].
L’importanza dei VA spinali nel drenaggio del FCS sembra supportata dall’osservazione del fatto che disturbare l’assorbimento cranico del FCS in seguito all’occlusione della cisterna magna con caolino abbia causato inizialmente un numero più che raddoppiato dell’ICP e della resistenza al deflusso (Ro). L’ICP, ma non il Ro, è ritornata a valori normali dopo sei settimane [84]. La valutazione istologica ha mostrato la formazione della siringa nel midollo cervicale e toracico, mentre le proteine ?con ?marker hanno lasciato lo spazio subaracnoideo spinale attraverso le radichette toraciche e lombo-sacrali. La presenza di un aumento continuo di Ro con un ICP normale dopo sei settimane ha indicato l’uso di vie di deflusso spinali perineurali per la compensazione dei disturbi di assorbimento cranico-FCS [84]. Nei ratti, utilizzando un metodo simile di ostruzione delle vie di assorbimento cranico, Voelz et al, mediante un tracciante alla ferritina, hanno dimostrato il passaggio dalla siringa del canale centrale attraverso le colonne rotte ependimali e dorsali nei vasi linfatici extradurali [85]. È stato suggerito che nell’essere umano in posizione verticale la pressione idrostatica differisca in modo significativo dalla pressione negli animali e, pertanto, le vie di deflusso spinali compensative del FCS possono anche essere di maggiore importanza. Va notato che mentre i tassi di pervietà del canale centrale negli esseri umani sono del 100% sotto 1 anno di età, ciò diminuisce sensibilmente a partire dalla seconda decade di vita. L’occlusione del canale centrale è iniziata a livello di T6 e L5 fino ad arrivare a S2 [86]. Questo percorso potenziale può non essere operativo dopo la seconda decade di vita.
Il fatto che i percorsi alternativi dopo uno dei siti di drenaggio del FCS siano anatomicamente alterati può essere riscontrato nell’ipovitaminosi A. È stato osservato in alcuni studi che sia una carenza che un eccesso di vitamina A potrebbero causare lo sviluppo d’idrocefalo nell’animale [31,87]. In questi studi, non vi era alcuna valutazione sulla funzione del percorso alternativo linfatico di assorbimento potenziale, che potrebbe essere stato compromesso in questi stessi animali. L’ipo e l’ipervitaminosi A si traducono in cambiamenti morfologici significativi dei VA. È stato riportato che i cambiamenti nella dura coinvolgono la porzione mucopolisaccaride e, dato che i VA sono ricchi di mucopolisaccaridi, potrebbe influenzare direttamente l’integrità di queste strutture e diminuirne le capacità funzionali [34,87,88]. L’assottigliamento del cappuccio cellulare dei VA e della fibrosi interstiziale è stato anche segnalato ed associato ad un aumento della resistenza al deflusso ed all’assorbimento del FCS [88,89]. Il fatto che l’aumento osservato della pressione del FCS non comporti lo sviluppo di idrocefalo suggerisce che il sistema linfatico possa rimanere intatto e fornire un percorso alternativo efficace per il drenaggio del FCS.
Dinamiche del drenaggio totale del FCS
Per valutare il rapporto tra la formazione e l’assorbimento del FCS, Davson ideò un sistema di infusione variabile che permetteva il controllo della velocità e della pressione di infusione [3]. Nel coniglio, la resistenza al deflusso del FCS ??era più di 10 volte maggiore di quella trovata negli umani. Questa differenza collima con il corrispondente tasso di formazione del FCS che nel coniglio è di 10-12 μl/min, mentre nell’uomo è di circa 350 μl/min. Egli concluse che se le pressioni del FCS in tutte le specie sono all’incirca le stesse, la resistenza al deflusso è allora inversamente proporzionale al volume del fluido che scorre attraverso lo spazio subaracnoideo. Cutler et al, [90] hanno condotto studi sui bambini utilizzando un sistema di perfusione ventricolo-cisternale per misurare il rapporto tra la formazione e l’assorbimento del FCS utilizzando una tecnica simile a quella descritta da Davson e altri [3]. Il calcolo della formazione e dell’assorbimento del FCS nelle persone era basato sulla diluizione dell’HSA radiomarcato in transito attraverso le vie ventricolo-spinali del FCS. La figura 12 mostra la curva derivata da questo studio. L’assorbimento o il deflusso del FCS (ml/min) è zero finché la pressione di scarico (pressione del FCS) supera 68 mm di FCS. Ciò è qualitativamente simile a quanto osservato nella perfusione dei VA isolati o a studi recenti sui flussi di liquidi in vivo ed in vitro misurati in una camera con una membrana di VA [5,24,25]. L’intercetto della riga di assorbimento e formazione attraversa ad un flusso di circa 0,37 ml/min e con una pressione di 112 millimetri di FCS. L’intercetto rappresenta l’uguaglianza tra la formazione e l’assorbimento del FCS. La pressione all’intercetto indica la pressione normale orizzontale a riposo in tali individui. L’assorbimento di FCS aumenta in modo lineare con l’innalzamento della pressione di deflusso del FCS entro una pressione del FCS di 68 – 250 mm, che è inoltre stata osservata in altri studi di infusione costante [90]. Questo studio ha anche rivelato che la formazione del FCS rimane invariata sulla stessa gamma di pressioni che è stata osservata anche nell’idrocefalo umano [3]. Al di là alla gamma di pressioni del FCS osservate in questo studio, la pressione venosa nel seno sagittale superiore è rimasta costante rispetto alla pressione del FCS. Nei bambini idrocefalici, Shulman e Ransohoff [91] hanno osservato che il consueto gradiente di pressione differenziale a favore del FCS al flusso del seno sagittale era stato negato. Nella stessa gamma di pressione, si è riscontrato che R0è resistente alla pressione, ma può deviare a pressioni notevolmente superiori [92]. Mann et al [93], utilizzando un sistema di infusione intraventricolare, hanno dimostrato che quando la velocità di infusione era pari o inferiore a 1,0 ml/min, l’innalzamento osservato della pressione del FCS raggiungeva uno stato stabile a quel tasso di livello di infusione. Ciò implica che il sistema di drenaggio o di deflusso del FCS era ancora operativo. Quando la velocità di infusione è stata aumentata a più di 1,0 ml/min circa, l’innalzamento della pressione che ne consegue non ha mai raggiunto una condizione stabile. In queste condizioni, la compliance del sistema venoso (riserva del volume) era esaurita e il sistema di deflusso è diventato inoperativo.
Si presume che la pressione del FCS a questa velocità di infusione abbia portato al collasso del sistema venoso cranico e alla perdita del differenziale di pressione FCS-venoso e la cessazione dell’assorbimento. La pressione a questo tasso d’infusione ha raggiunto un valore > 900 mm H20. Un metodo utile per valutare la compliance, l’assorbimento e la formazione del FCS è l’iniezione di bolo [41]. Questo metodo alternativo esprime la relazione tra pressione-volume come un indice di volume della pressione (PVI), ovvero il volume necessario per aumentare la pressione del FCS di 10 volte. La compliance (C) può essere calcolata utilizzando una singola iniezione di bolo e registrando il volume iniettato (V) e la pressione alla base (Po) e al picco (Pp) come V/log10 Pp/Po. Questo metodo del bolo consente non solo il calcolo della compliance (e l’inverso, ovvero l’elastanza) del sistema del FCS, ma anche la resistenza al deflusso dei percorsi assorbenti [41]. È stato determinato nel gatto che circa i 2/3 della compliance erano nel compartimento cranico (il 48% nel sopratentoriale ed il 20% nell’infratentoriale) e 1/3 nel compartimento spinale [41]. Il compartimento cranico contava circa l’84% dell’assorbimento totale di FCS in condizioni normali. La capacità di assorbimento spinale del 16% dell’assorbimento totale suggerisce un ruolo relativamente minore dell’aracnoide spinale per compensare le necessità di accumulo di volume all’interno dello spazio del FCS [42]. Lofgren e Swetnow hanno dimostrato un cambiamento notevole del volume di distribuzione tra i compartimenti cranici e spinali mano a mano che il volume del FCS aumentava oltre il valore normale, il che ha portato a un rovesciamento della distribuzione dell’aumento del volume in favore del compartimento spinale [94]. L’analisi degli studi sull’uomo di Katzman e Hussey [95] effettuata da parte di Marmarou et al [41] ha rivelato che il PVI totale nell’uomo è di circa 25 ml e la distribuzione della compliance tra i compartimenti cerebrali e spinali è di 2 a 1.
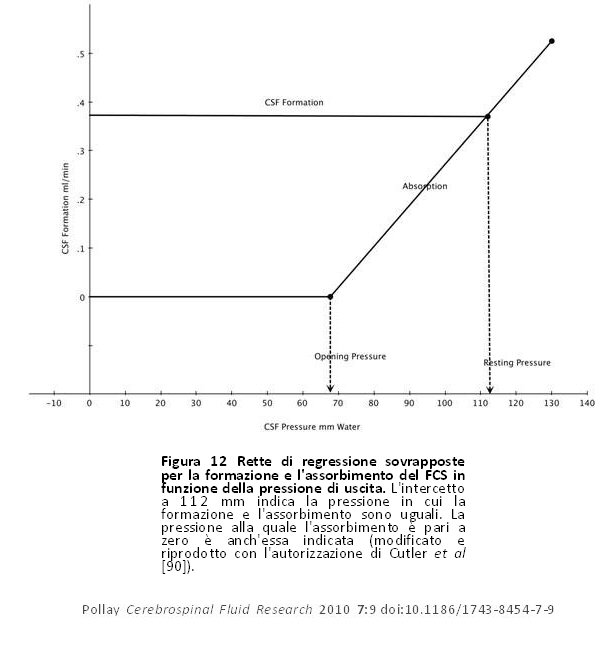 Figura 12 Rette di regressione sovrapposte per la formazione e l’assorbimento del FCS in funzione della pressione di uscita. L’intercetto a 112 mm indica la pressione in cui la formazione e l’assorbimento sono uguali. La pressione alla quale l’assorbimento è pari a zero è anch’essa indicata (modificato e riprodotto con l’autorizzazione di Cutler et al [90]). Figura 12 Rette di regressione sovrapposte per la formazione e l’assorbimento del FCS in funzione della pressione di uscita. L’intercetto a 112 mm indica la pressione in cui la formazione e l’assorbimento sono uguali. La pressione alla quale l’assorbimento è pari a zero è anch’essa indicata (modificato e riprodotto con l’autorizzazione di Cutler et al [90]). |
Ekstedt [96] scoprì che il rapporto tra il deflusso e la pressione del FCS era rettilineo fino ad una pressione del FCS di circa 6 kPa (61 cm H20) dopo aver raggiunto una pressione del FCS di circa 60 mm H20 necessaria per iniziare il deflusso del FCS. Ciò gli suggerì che una volta che i VA sono aperti non possono essere più dilatati dalla pressione. Risulta chiaro ora che la resistenza al deflusso e la conduttanza che misurò nell’uomo rappresentano sia il deflusso attraverso i VA cranici e spinali sia quello attraverso il sistema linfatico perineurale. Ciò ha suggerito che i valori di resistenza al deflusso del sistema totale (Ro) ed il percorso della capacità di assorbimento possono rappresentare un valore medio combinato per entrambi i percorsi. Utilizzando la tecnica dell’iniezione di bolo, Sokolowski [97] costruì una serie di curve pressione-tempo (PT) da cui potevano essere derivati sia un piano pressione-volume (PV) che un piano volume-tempo (VT). Il piano PV rappresenta il grado di distensibilità del sistema e la disponibilità dello spazio di riserva. Maggiore è la pressione, più veloce sarà il drenaggio del liquido in eccesso dal sistema. Le curve calcolate volume-tempo (VT) mostrano che il tasso di assorbimento è una funzione della pressione. La pendenza di questo tipo di curva si presume possa essere determinata principalmente applicando il meccanismo di drenaggio, anche se il grado di elasticità nel sistema di compliance è anch’esso un fattore da considerare. La curva monoesponenziale osservata rappresenta il sistema normale in cui l’assorbimento del FCS è rapido e completo. Quando il/i meccanismo/i di assorbimento si scompongono (come nel paziente idrocefalico), si nota una curva bi-compartimentale che può indicare un assorbimento difettoso, prolungato ed incompleto. Ciò suggerì a Sokolowski che l’assorbimento globale del FCS fosse composto da due meccanismi di deflusso: un sistema di assorbimento ad alta pressione e un altro a bassa pressione che operano in armonia in condizioni normali [97]. In alternativa, ciò può anche rappresentare un sistema secondario per l’assorbimento in presenza di un difetto di assorbimento primario. Nelson e Goodman hanno sostenuto questo concetto e proposto che il sistema del FCS fosse composto da una serie di meccanismi valvolari con una soglia di apertura progressivamente più alta ed elementi inferiori o superiori bloccati selettivamente [98]. Ciò è in sostanziale accordo con quanto riportato da Lorenzo et al [99] che studiarono la relazione tra l’assorbimento del FCS e la pressione di deflusso nell’idrocefalo umano. Essi hanno osservato un difetto di assorbimento di tipo 1 che consisteva in una pressione di apertura > 68 mmH2O, ma in un tasso di assorbimento di fluido di 0,0069 ml/min/mm che era simile al valore di controllo di 0,0076 ml/min/mm. Il difetto di tipo 2 aveva una pressione di apertura normale, ma il tasso di assorbimento era ben al di sotto del valore di controllo con 0,0026 ml/min/mm. Un altro approccio menzionato in precedenza è il modello ovino in cui le caratteristiche di deflusso dei sistemi di VA e linfatico sono state valutate separatamente per raccolta o isolamento di deflusso singolo a causa di un’ostruzione fisica [68-70,100]. I metodi di raccolta isolati hanno indicato che entrambi i sistemi contavano circa la metà dell’assorbimento del FCS. In media, un aumento dell’ICP di 10 cmH2O innalzava la clearance del FCS dei VA e linfatico rispettivamente di 2,7 e 3,9 volte. Studi più datati sugli ovini maturi [66,67] rivelano che il drenaggio del FCS attraverso il sistema linfatico rappresenta il 40-48% del totale. Negli animali giovani, il sistema di deflusso linfatico sembra essere dominante in larga misura dallo sviluppo un po’ ritardato dei VA sia per quanto riguarda il numero che la maturità [14,79]. Il blocco del percorso perineurale olfattivo alla lamina cribiforme ha compromesso notevolmente il trasporto del FCS dimostrando l’importanza del sistema linfatico nel drenaggio totale del FCS.Nella figura 13 è chiaro che l’assorbimento del FCS è significativamente ridotto dopo il blocco del deflusso del FCS attraverso la lamina cribiforme in un animale a cui è stato negato l’accesso al compartimento spinale [72]. In questo preparato sperimentale, la curva della velocità di deflusso-ICP si è spostata verso destra e l’aumento della resistenza al deflusso ha richiesto una pressione maggiore per ottenere lo stesso flusso osservato prima dell’ostruzione. I valori dell’ICP sono quelli al di sopra della pressione di apertura. Tale studio suggerisce che i VA cranici rappresentano un sistema di deflusso operante a pressioni elevate di FCS rispetto a quella del flusso attraverso la lamina cribiforme nel sistema di deflusso linfatico nasale. Mollanji et al [100] hanno bloccato la lamina cribiforme e in alcuni esperimenti hanno anche isolato il compartimento spinale negli ovini e testato il sistema con infusioni a flusso o a pressione costanti. La percentuale del trasporto totale del FCS a varie pressioni superiori alla pressione di apertura è riportata nella figura 14.
Si presume, che nelle pecore, la pressione di apertura (OP) per il flusso del FCS sia simile a quella osservata nell’uomo (comunicazione personale-Miles Johnston) e questo sembra proprio essere quanto avviene in questo studio. Dalla figura 14A e 14B, il deflusso comincia a verificarsi più o meno al contempo sopra alla pressione di apertura, anche se la pressione necessaria affinché il flusso non-linfatico (villi cranici e spinali) eguagli il 50% del trasporto totale del FCS è quasi 10 cm al di sopra della OP. Quando il compartimento spinale viene isolato mediante ligazione (Figura 14B), il punto in cui i sistemi dei VA (cranici) e linfatici raggiungono il 50% del trasporto totale del FCS si trova a un ICP più elevato e suggerisce l’importanza del sistema dei VA spinali nell’assorbimento totale del FCS. Ciò non è affatto sorprendente dato che il compartimento spinale ha dimostrato di essere responsabile fino al 25% circa del sistema di drenaggio totale del FCS.
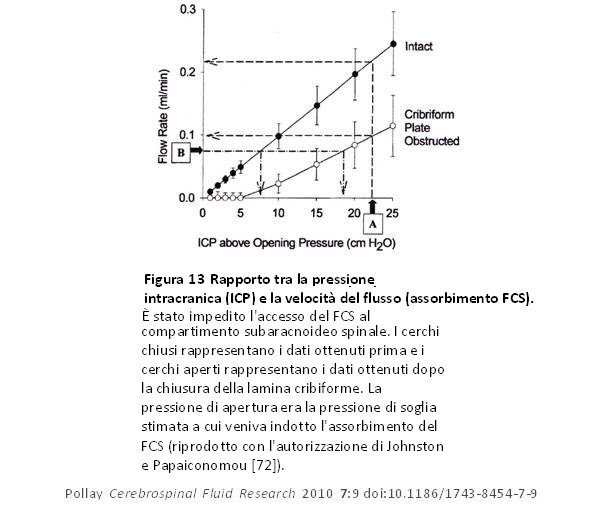 Figura 13 Rapporto tra la pressione intracranica (ICP) e la velocità del flusso (assorbimento FCS). È stato impedito l’accesso del FCS al compartimento subaracnoideo spinale. I cerchi chiusi rappresentano i dati ottenuti prima e i cerchi aperti rappresentano i dati ottenuti dopo la chiusura della lamina cribiforme. La pressione di apertura era la pressione di soglia stimata a cui veniva indotto l’assorbimento del FCS (riprodotto con l’autorizzazione di Johnston e Papaiconomou [72]). Figura 13 Rapporto tra la pressione intracranica (ICP) e la velocità del flusso (assorbimento FCS). È stato impedito l’accesso del FCS al compartimento subaracnoideo spinale. I cerchi chiusi rappresentano i dati ottenuti prima e i cerchi aperti rappresentano i dati ottenuti dopo la chiusura della lamina cribiforme. La pressione di apertura era la pressione di soglia stimata a cui veniva indotto l’assorbimento del FCS (riprodotto con l’autorizzazione di Johnston e Papaiconomou [72]). |
 Figura 14 Stime sulla proporzione del trasporto totale del FCS attraverso la lamina cribiforme (cerchi pieni) e gli altri percorsi (villi aracnoidei cranici e spinali) con cerchi aperti in esperimenti a flusso (A) e pressione (B) costanti. Il drenaggio cribiforme è il sito dominante a pressioni ICP basse e moderate. In A, il compartimento spinale è intatto, mentre in B il compartimento spinale è bloccato (riprodotto con l’autorizzazione di Mollanji et al [100]). Figura 14 Stime sulla proporzione del trasporto totale del FCS attraverso la lamina cribiforme (cerchi pieni) e gli altri percorsi (villi aracnoidei cranici e spinali) con cerchi aperti in esperimenti a flusso (A) e pressione (B) costanti. Il drenaggio cribiforme è il sito dominante a pressioni ICP basse e moderate. In A, il compartimento spinale è intatto, mentre in B il compartimento spinale è bloccato (riprodotto con l’autorizzazione di Mollanji et al [100]). |
La capacità massima dei sistemi di drenaggio del FCS nel gestire il volume di liquor prodotto prima di sviluppare la ventricolomegalia non è stata completamente definita. La misura della formazione del FCS mediante drenaggio esterno è variata notevolmente in pazienti con papilloma del plesso coroideo con idrocefalo associato. In alcuni casi, la velocità di formazione così determinata non era molto diversa rispetto al normale intervallo di valori. Tuttavia, non vi era alcuna valutazione della capacità di assorbimento del sistema di deflusso misurata allo stesso tempo del drenaggio. Ciò indica che il sistema di deflusso potrebbe inoltre essere negativamente influenzato in tali pazienti]. La risposta del sistema di FCS all’innalzamento dei tassi di formazione del FCS (sovrapproduzione) è stata studiata in un paziente con idrocefalo associato a un papilloma del plesso coroideo da Eisenberg et al [101], mediante un sistema di perfusione ventricolo-lombare. Hanno dimostrato che la capacità di assorbimento del sistema di deflusso totale supera di quattro volte (1,43 ml/min) il tasso di formazione del FCS nei bambini (normale ~ 0,35 ml/min). Il tasso di assorbimento (Va) in tal caso era stato misurato a 130 mmH2O e rilevato essere 0,59 ml/min, ovvero un valore quasi uguale a quello trovato in bambini normali con percorsi non ostruiti (Va = 0.61 ml/min). Dopo la rimozione del tumore del plesso coroideo, la pressione intracranica nel bambino è ritornata normale e la dimensione della testa è scesa al 50°percentile d’età. Si può concludere da questo studio che sopra il normale livello massimo di assorbimento di circa 1,0 ml/min, il fallimento della compliance cerebrale nell’accogliere il fluido in eccesso e l’effetto della pressione del FCS sul sistema venoso cranico comporterà un accumulo di liquidi e la ventricolomegalia. Ciò si dimostra in linea con gli studi di perfusione di Mann effettuati sui cani che ha dimostrato che la capacità di assorbimento del sistema del FCS viene superata quando l’infusione nello spazio subaracnoideo sopracorticale è > 1,0 ml/min [93].
Conclusioni
Le prove finora disponibili confermano l’importanza sia del sistema dei villi aracnoidei cranici (VA) che di quello di drenaggio linfatico nell’uscita del liquor dallo spazio subaracnoideo. I VA spinali hanno un ruolo minore, anche se importante, soprattutto in presenza di insufficienza dei sistemi primari. L’anatomia di entrambi i sistemi conferma la tesi secondo la quale entrambi agiscono come un percorso unidirezionale aperto tra il FCS subaracnoideo e il sistema circolatorio vascolare. Il sistema di deflusso linfatico avviene principalmente attraverso lo spazio perineurale del nervo olfattivo che attraversa la lamina cribiforme. Altri nervi cranici sono stati implicati, ma non ci sono prove a supporto di un loro ruolo importante nel drenaggio liquorale. Il sistema aracnoideo cranico sembra permettere il movimento del FCS attraverso i vacuoli intracellulari e le schisi intercellulari. Nei primati si trovano principalmente lungo la dura del seno sagittale superiore vicino all’ingresso della vena occipito-parietale, mentre negli animali inferiori lungo il seno trasverso vicino alla confluenza dei seni. Si è dimostrato che il sistema linfatico si sviluppa prima di quello dei VA e sembra pertanto un percorso di deflusso del FCS dominante nel tardo periodo fetale e nel primo periodo neonatale. Ci sono prove convincenti del fatto che il sistema dei VA perda efficacia con l’età, cosa che può influenzare il tasso di ricambio totale del FCS, con possibili conseguenze neurodegenerative. La questione inerente il fatto che l’ostruzione di uno dei sistemi possa portare da sola ad idrocefalo è ancora aperta poiché ci sono poche prove convincenti disponibili che separano lo stato dei singoli componenti in un ambiente clinico patologico.
Il sistema di drenaggio globale del FCS dei primati diventa operativo a circa 50 a 70 mm di pressione del FCS e normalmente fluisce al di fuori del sistema subaracnoideo ad un gradiente di pressione differenziale da 3 a 4 mm Hg. La velocità del flusso normale non varia con l’età, ma la media è di circa 0,35 ml/min. La capacità massima del sistema di drenaggio globale sembra essere di circa 1,0 ml/min, valore al di sopra del quale il liquido si accumula nel sistema ventricolare sotto l’aumento della pressione. Non è possibile determinare nei primati la percentuale del trasporto totale di FCS dovuta ai sistemi linfatico e di deflusso dei VA, anche se è stato osservato nell’uomo che vi è un sistema di deflusso sia ad alta che a bassa pressione. È forse ragionevole ritenere, sulla base degli studi sugli ovini, che il sistema linfatico perineurale olfattivo è il sistema a pressione inferiore. Non ci sono dati sperimentali che indichino con certezza la proporzione di drenaggio liquorale totale dovuta ad uno dei due sistemi con pressione a riposo del FCS nei primati.
Conflitto d’interessi
Gli autori dichiarano che non ci sono conflitti d’interessi.
Contributi degli autori
L’autore Michael Pollay è l’unica persona interamente responsabile del contenuto di questo manoscritto. La traduzione in italiano è stata curata da Osteopatia News rispettando i Creative Commons.
Riferimenti bibliografici
- Brunori A, Vagnozzi R, Giuffre R Antonio Pacchioni (1665-1726) early studies of the dura mater. J Neurosurg 1993, 78:515-518.
- Trolard D: Les Lacunes Veineuses de la dura-mère. J de Lanatomie 1892, 38:28-56.
- Davson H, Segal MB: Physiology of the Cerebrospinal Fluid and Blood-Brain Barriers London: CRC Press; 1996.
- Key A, Retzius G: Studien in derAnatomie des Nervensystems und des Bindegewebes Stockholm: Samsin and Wallen; 1875.
- Welch K, Friedman V: The Cerebrospinal fluid valves. Brain 1960, 83:454-469.
- Davson H: Physiology ofthe ocular and cerebrospinal fluids London: Churchill; 1956.
- Elman R: Spinal arachnoid granulations with special reference to the CSF. Bull Johns Hopkins Hosp 1923, 34:99-104.
- Wislocki GB: The cytology of the cerebrospinal fluid pathways. In Special Cytology Volume III. 3rd edition. Edited by: Cowdry EV. New York: Hoeber; 1932.
- Welch K, Pollay M: Perfusion of particles through the arachnoid villi of the monkey. Am J Physiol 1961, 201:651-654.
- Pollay M, Welch K: The function and structure of the canine arachnoid villi. JSR 1962, 2:307-312.
- Clark WE, Legros : On the Pacchionian bodies. JAnat 1920, 55:40-48.
- Yamashita K, Morita F, Otsuki Y, Magari S: Morphology of arachnoid granulations and villi in the region of the sella tursica-light microscopy and three-dimensional image analysis. Okajima Folia AnatJpn 1989, 66:81-97.
- Gomez DG, Ehrmann JE, Potts DG, Pavese AM, Gilanian A: The arachnoid granulations of the newborn human an ultrastructural study. Int J Dev Neurosci 1983, 1:139-147.
- Papaiconomou C, Bozanovic-Sosic R, Zakharov A, Johnston M: Does neonatal cerebrospinal fluid absorption occur via arachnoid projections or extracranial lymphatics? Am J Physiol Regul Integr Comp Physiol 2002, 283:R869-R876.
- Johanson CE, Duncan JA III, Klinge PM, Brinker T, Stopa EG, Silverberg GD: Multiplicity of cerebrospinal fluid functions: New challenges in health and disease. Cerebrospinal Fluid Res 2008, 5:10.
- Rubenstein E: Relationship of senescence of cerebrospinal fluid circulatory system to dementia of the aged. Lancet 1998, 24:283-285.
- Shabo AL, Brightman MW: Permeability to peroxidase of the endothelium in the arachnoid villus of the monkey. Anat Rec 1972,172:404.
- Gomez DG, Potts DG, Deonarine V, Reilly DF: Effect of pressure gradient changes on the morphology of arachnoid villi and granulations of the monkey. Lab Invest 1973, 28:648-657.
- Tripathi RC: Tracing the bulk outflow route of cerebrospinal fluid by transmission and scanning electron microscopy. Brain Res 1974, 80:503-506.
- Tripathi RC: Ultrastructure of Schlemm’s canal in relation to aqueous outflow. Exp Eye Res 1968, 7:335-341.
- Levine JE, Povlishock DP, Becker DP: The morphological correlates of primate cerebrospinal fluid absorption. Brain Res 1982, 24:31-41.
- Upton ML, Weller RO: The morphology of cerebrospinal fluid drainage pathways in human arachnoid granulations. J Neurosurg 1985, 63:867-875.
- d’Avella D, Cicciarello R, Albiero F, Andrioli G: Scanning electron microscopic study of human arachnoid villi. J Neurosurg 1983, 59:620-626.
- Grzybowski DM, Holman DW, Katz SE, Lubow M: In vitro model of cerebrospinal fluid outflow through human arachnoid granulations. Invest Ophthalmol Vis Sci 2006, 47:3664-3672.
- Glimcher SA, Holman DW, Lubow M, Grzybowski DM: Ex vivo model of cerebrospinal fluid outflow across human arachnoid granulations. Invest Ophthalmol Vis Sci 2008, 49:4721 -4728.
- Shulman K, Yarnell P, Ransohoff J: Dural sinus pressure. Arch Neurol 1964, 10:575-580.
- Mann JD, Butler AB, Johnson RN, Bass NH: Clearance of macromolecular and particulate substances from the cerebrospinal fluid system of the rat. J Neurosurg 1979, 50:343-348.
- McComb JG, Davson H, Hyman S, Weiss MH: Cerebrospinal fluid drainage as influenced by ventricular pressure in the rabbit. J Neurosurg 1982, 56:790-797.
- Massicotte EM, Del Bigio MR: Human arachnoid villi response to subarachnoid hemorrhage: possible relationship to chronic hydrocephalus. J Neurosurg 1999, 91:80-84.
- Gutierrez Y, Friede RL, Kaliney WJ: Agenesis of arachnoid granulations and its relationship to communicating hydrocephalus. Neurosurg 1975,43:553-558.
- Millen JW, Woollam DHM: The effect of the duration of Vitamin A deficiency in female rabbits upon the incidence of hydrocephalus in their young. J Neurol Neurosurg Psychiatr 1956, 19:17-20.
- Calhoun MC, Hurt HD, Eaton HD, Rousseau JE Jr, Hall RC Jr: Rate of formation and absorption of cerebrospinal fluid in bovine hypovitaminosis A. J Dairy Sci 1967, 50:1486-1494.
- Abernathy RS: Bulging fontanelle as presenting sign in cystic fibrosis. Vitamin A metabolism and effect of cerebrospinal fluid pressure. Am J Dis Child 1976, 130:1360-1362.
- Gorgacz EJ, Nielsen SW, Frier HI, Eaton HD, Rousseau JE Jr: Morphological alterations associated with decreased cerebrospinal fluid pressure in chronic bovine hypervitaminosis A. Am J Vet Res 1975, 36:171-180.
- Tabassi A, Salmasi AH, Jalali M: Serum and cerebrospinal fluid vitamin A concentrations in idiopathic intracranial hypertension. Neurology 2005, 64:1893-1896.
- Brierley JB, Fields EJ: The connection of the spinal subarachnoid space with the lymphatics. J Anat 1948, 82:153-166.
- Welch K, Pollay M: The spinal arachnoid villi of the monkeys Ceropithe aethiops and Macaca irus. Anat Rec 1963, 145:43-48.
- Gomez DG, Chambers AA, Di Benedetti AT, Potts DG: The spinal cerebrospinal fluid pathways. Neuroradiol 1974, 8:61-66.
- Kido DK, Gomez DG, Pavese AM, Potts DG: Human spinal arachnoid granulations. Neuroradiol 1976, 11:221-228.
- Tubbs RS, Hansasuta A, Stetler W, Kelly DR, Blevins D, Humphrey R, Chua GD, Shoja MM, Loukas M, Oakes WJ: Human spinal arachnoid villi revisited: immunohistological study and review of the literature. J Neurosurg Spine 2007, 7:328-331.
- Marmarou A, Shulman K, LaMorgese J: Compartmental analysis of compliance and outflow resistance of the cerebrospinal fluid system. J Neurosurg 1975, 43:523-534.
- Bozanovic-Sosic R, Molianji R, Johnston M: Spinal and cranial contributions to total cerebrospinal fluid transport. J Physiol Regul Integr Comp Physiol 2001, 281 :R909-916.
- Edsbagge M, Tissel M, Jacobsson L, Wikkelso C: Spinal cerebrospinal fluid absorption in healthy individuals. Am J Physiol Regul Integr Comp Physiol 2004, 287:R1450-R1455.
- Koh L, Zakharov A, Johnston M: Integration of the subarachnoid space and lymphatics: Is it time to embrace a new concept of cerebrospinal fluid absorption. Cerebrospinal Fluid Res 2005, 2:6.
- Mortensen OA, Sullivan WE: The Cerebrospinal fluid and the cervical lymph nodes. Anat Rec 1933, 56:359-362.
- Faber WM: The nasal mucosa and the subarachnoid space. Am J Anat 1937, 62:121-148.
- Czerniawska A: Experimental investigation on the penetration of 198 AU from nasal mucous membrane into cerebrospinal fluid. Acta Otolaryngol 1970, 70:58-61.
- Jackson RT, Tigges J, Arnold W: Subarachnoid space of the central nervous system, nasal mucosa and lymphatic system. Arch Otolaryngol 1979, 105:180-184.
- Courtice FC, Simmonds WJ: The removal of protein from the subarachnoid space. Austral J Exper Biol Med Sci 1951, 29:255-263.
- Simmonds WJ: The absorption of labeled erythrocytes from the sub arachnoid space of the rabbit. J Exper Biol Med Sci 1953, 31:77-83.
- Cserr HF, Harling-Berg CJ, Knopf PM: Drainage of brain extracellular fluid into blood and deep cervical lymph and its immunological significance. Brain Pathol 1992, 4:269-276.
- Cserr HF, DePasquale M, Herling-Berg CJ, Park JT, Knopf PM: Afferent and efferent arms of the tumoral immune response to CSF-administered albumin in a rat model with normal blood-brain barrier permeability. J Neuroimmunol 1992, 41:195-202.
- Weller RO, Kida S, Zhang ET: Pathways of fluid drainage from the brain-morphological aspects and immunological significance in rat and man. Brain Pathol 1992, 2:277-284.
- Ludemann W, Berens von Rautenfeld D, Samii M, Brinker T: Ultrastructure of the CSF fluid outflow along the optic nerve into the lymphatic system. Childs NervSyst 2005, 21:96-103.
- Manzo RP, Gomez DG, Potts DG: Cerebrospinal fluid absorption in the rabbit Inner ear pathways. Acta Otolaryngol 1990, 109:389-396.
- Bradbury MWB, Cole DF: The role of the lymphatic system in drainage of cerebrospinal fluid and the aqueous humor. J Physiol 1980, 299:353-365.
- Bradbury MWB, Cserr HF, Westrop RJ: Drainage of cerebral interstitial fluid into deep cervical lymph of the rabbit. Am J Physiol 1981, 240:F329-F336.
- Bradbury MW, Westrop RJ: Factors influencing exit of substances from cerebrospinal fluid into deep cervical lymphatics of the sheep. J Physiol 1983, 339:519-534.
- Gomez DG, Fenstermacher JD, Manzo RP, Johnson D, Potts DG: Cerebrospinal fluid absorption in the rabbit: olfactory pathways. Acta Otolaryngol 1985, 100:429-436.
- Lowhagen P, Johnansson BB, Nordborg C: The nasal route of cerebrospinal fluid drainage in man. A light microscope study. Neuropathol Appl Neurobiol 1994, 20:543-550.
- Kida S, Pentazis A, Weller RO: Cerebrospinal fluid drains directly from the subarachnoid space into nasal lymphatics in the rat. Anatomy, histology and immunological significance. Neuropathol Appl Neurobiol 1993, 19:480-488.
- Gomez DG, Manzo RP, Fenstermacher JD, Potts DG: Cerebrospinal fluid absorption in the rabbit. Optic pathways. Graefes Arch Exp Ophthalmol 1988, 226:1-7.
- Leeds SE, Kong AK, Wise BL: Alternate pathways for drainage of cerebrospinal fluid in the canine brain. Lymphology 1989, 22:144-146.
- Johnston M, Zakharov A, Papaiconomou C, Salmasi G, Armstrong D: Evidence of connections between cerebrospinal fluid and nasal lymphatic vessels in humans, non-human primates, and other mammalian species. Cerebrospinal Fluid Res 2004, 1:2.
- Kida S, Pantazis A, Weller RO: CSF drains directly from the subarachnoid space into nasal lymphatics in the rat. Anatomy, histology and immunological significance. Neuropathol Appl Neurobiol 1993,19:480-488.
- Boulton M, Young A, Hay J, Armstrong D, Flessner M, Schwartz M, Johnston M: Drainage of cerebrospinal fluid through lymphatic pathways and arachnoid villi in sheep: measurement of 125 I- albumin clearance. Neuropathol Appl Neurobiol 1996, 22:325-333.
- Boulton M, Flessner M, Armstrong D, Hay J, Johnston M: Determination of volumetric CSF absorption into extracranial lymphatics in sheep. Am J Physiol 1998, 274:R88-R96.
- Boulton M, Armstrong D, Flessner M, Hay J, Szalai JP, Johnston M: Raised intracranial pressure increases cerebrospinal fluid drainage through arachnoid villi and extracranial lymphatics. Am J Physiol 1998, 275:R889-896.
- Boulton M, Flessner M, Armstrong D, Mohamed R, Hay J, Johnston M: Contribution of extracranial lymphatics and arachnoid villi to the clearance of a cerebrospinal fluid transport in the rat. Am J Physiol 1999, 276:R818-R823.
- Silver J, Kim C, Mollanji R, Johnston M: Cerebrospinal fluid outflow resistance in sheep: impact of blocking cerebrospinal fluid transport through the cribiform plate. Neuropathol Appl Neurobiol 2002, 28:67-74.
- Papaiconomou C, Zakharov A, Azzizi N, Djenic j, Johnston M: Reassessment of the pathways responsible for cerebrospinal fluid absorption in the neonate. Childs NervSyst 2004, 20:29-36.
- Johnston M, Papaiconomou C: Cerebrospinal fluid transport; a lymphatic perspective. News Physiol Sci 2002, 17:227-230.
- Koh L, Zakharov A, Nagra G, Armstrong D, Friendship R, Johnston M: Development of cerebrospinal fluid absorption sites in the pig and rat: connections between the subarachnoid space and lymphatic vessels in the olfactory turbinates. Anat Embryol (Berl) 2006, 211:335-344.
- Mollanji R, Papaiconomou C, Boulton M, Midha R, Johnston M: Comparison of CSF transport in fetal and adult sheep. Am J Physiol Regul Integr Comp Physiol 2001, 281:R1215-R1223.
- Jones HC: Cerebrospinal fluid pressure and resistance to absorption during development in normal and mutant mice. Exp Neurol 1985, 90:162-172.
- Jones HC, Deane R, Bucknall RM: Development changes in cerebrospinal fluid pressure and resistance to absorption in rats. Dev Brain Res 1987, 33:23-30.
- Nagra G, Johnston MG: Impact of ageing on lymphatic cerebrospinal fluid absorption in the rat. Neuropathol Appl Neurobiol 2007, 33:684-691.
- Nagra G, Li J, McAllister JP II, Miller J, Wagshul M, Johnston M: Impaired lymphatic cerebrospinal fluid absorption in a rat model of kaolin-induced communicating hydrocephalus. Am J Physiol Regul Integr Comp Physiol 2008, 94:R1752-1759.
- Rammling M, Madan M, Paul L, Behnam B, Pattisapu JV: Evidence for reduced lymphatic cerebrospinal fluid absorption in the H-Tx rat hydrocephalus model. Cerebrospinal Fluid Res 2008, 5:15.
- Macauley N, Zeuthen T: Water transport between central nervous system compartments: contributions of aquaporins and co transporters. Neuroscience 2009, 14:S1873-7544.
- Tourdias T, Dragonu I, Fushimi Y, Deloire MS, Boiziau C, Brochet B, Moonen C, Petry KG, Dousset V: Aquaporin 4 correlates with apparent diffusion coefficient and hydrocephalus severity in the rat brain: a combines MRI histological study. Neuroimage 2009, 47:659-666.
- Shen XQ, Miyajima M, Ogino I, Arai H: Expression of the water channel protein aquaporins 4 in the H-Tx rat: possible compensatory role in spontaneously arrested hydrocephalus. J Neurosurg 2006, 195:459-464.
- Duong DH, O’Malley S, Sekhar LN, Wright DG: Postoperative Hydrocephalus in cranial base surgery. Skull Base Surgery 2000,10:197-200.
- Ludemann W, Kondziella D, Tienken K, Brinker T: Spinal cerebrospinal fluid pathways and their significance for the compensation of kaolin-hydrocephalus. Acta Neurochir Suppl 2002, 81:271-273.
- Voelz K, Kondziella D, Beren von Rautenfeld DB, Brinker T, Luedemann W: A ferritin tracer study of compensatory spinal cerebrospinal fluid outflow pathways in kaolin-induced hydrocephalus. Acta Neuropath 2007, 113:569-575.
- Yasui K, Hashizume Y, Yoshida M, Kameyama T, Sobue G: Age related morphological changes of the central canal of the human spinal cord. Acta Neuropath 1999, 97:253-259.
- Van der Lugt JJ, Prozesky L: The pathology of blindness in new born calves caused by hypovitaminosis A. Onderstepoort J Vet Res 1989, 56:99-109.
- Cousins RJ, Eaton HD, Rousseau JE jr, Hall RC jr: Biochemical constituents of the dura mater in vitamin A deficiency. J Nutrition 1969, 97:409-418.
- Gorgacz EJ, Nielsen SW, Frier HI, Eaton HD, Rousseau JE Jr: Morphological alterations associated with decreased cerebrospinal fluid pressure in chronic bovine hypervitaminosis A. Am J Vet Res 1975, 36:171-18.
- Cutler RWP, Page L, Galicich J, Watters GV: Formation and absorption of cerebrospinal fluid in man. Brain 1968, 91:707-720.
- Shulman K, Ransohoff J: Sagittal sinus pressure in hydrocephalus. J Neurosurg 1965, 23:169-173.
- Andersson N, Malm J, Eklund A: Dependency of cerebrospinal fluid outflow resistance on intracranial pressure. J Neurosurg 2008, 109:918-922.
- Mann JD, Maffeo CJ, Rosenthal JD, Johnson RN, Butler AB, Bass NH: Regulation of intracranial pressure in rat, dog and man: manometric assessment of cerebrospinal fluid dynamics using a constant flow perfusion system. Trans Am Neurol Assoc 1976, 101:182-185.
- Lofgren J, Swetnow NN: Cranial and spinal components of the cerebrospinal fluid pressure volume curve. Acta NeurolScand 1973,49:575-585.
- Katzman R, Hussey F: A simple constant infusion manometric test for the measurement of cerebrospinal fluid absorption. 1. Rational and method. Neurology 1970, 20:534-544.
- Ekstedt J: Cerebrospinal fluid hydrodynamic studies in man. J Neurol Neurosurg Psychiat 1977, 40:105-119.
- Sokolowski SJ: Bolus injection test for measurement of cerebrospinal fluid absorption. J Neurol Sci 1976, 28:491-504.
- Nelson JR, Goodman SJ: An evaluation of the cerebrospinal fluid infusion test for hydrocephalus. Neurology 1971, 21:1037-1053.
- Lorenzo AV, Page LK, Watters GV: Relationship between cerebrospinal fluid formation, absorption and pressure in human hydrocephalus. Brain 1970, 93:679-690.
- Mollanji R, Bozananovic-Sosic I, Silver B LI, Kim C, Midha R, Johnston M: Intracranial pressure accommodation is impaired by blocking pathways leading to extracranial lymphatics. Am J Physiol Regulatory Integrative Comp Physiol 2001, 280:R1573-R1581.
- Eisenberg HM, McComb JG, Lorenzo AV: Cerebrospinal fluid overproduction and hydrocephalus associated with choroid plexus papilloma. J Neurosurg 1974, 40:381-385.