
Lo scambio di segnali tra i sistemi immunitario, endocrino e nervoso
per l’immunoregolazione e le funzioni cerebrali
Hugo O. Besedovsky, Adriana del Rey
Questo articolo offre una visione personale su come il concetto dell’esistenza di una rete di interazioni immuno-neuro-endocrine si è evoluto negli ultimi 30 anni. L’argomento principale esaminato è l’importanza dello scambio di segnali tra i sistemi immunitario, endocrino e nervoso per l’immunoregolazione e le funzioni cerebrali. Particolare enfasi è stata data ai circuiti che interessano i prodotti della cellula immunitaria, l’asse ipotalamo- ipofisi-surrene ed il sistema nervoso simpatico. L’attività di questi circuiti può interessare le funzioni immunitarie e il decorso delle malattie infiammatorie, autoimmuni e infettive. E’ discussa anche l’evidenza sempre più notevole del fatto che le citochine di origine cerebrale giocano un ruolo importante nella fisiologia del cervello e nell’integrazione della rete immuno-neuro-endocrina.
Scarica la sintesi in italiano in formato PDF (236 Kb)
L’articolo originale (Brain, Behavior, and Immunity 2007; 21: 34–44)
1. INTRODUZIONE: ASPETTI GENERALI E DEFINIZIONI
Non molto tempo fa (almeno per quanto riguarda la misura del tempo nella scienza), quando si intendeva introdurre un articolo sulle interazioni tra i sistemi immunitario, endocrino e nervoso, ci si riferiva ad un campo “emergente”. Questo tipo di introduzione non è più giustificabile, dato che quest’area di ricerca è ora decisamente consolidata. La comparsa di “Brain, Behavior and Immunity” 20 anni fa, è stata un pilastro importante
per il suo consolidamento. Prima era difficile convincere recensori delle riviste focalizzate su una specifica specializzazione che i confini tra le discipline sono relativi e talvolta auto- imposti. Gli autori sostengono che la serie speciale di BBI (Brain,Behavior and Immunity) sia importante perché fornisce una prospettiva storica che potrebbe servire a influenzare correnti di pensiero e sviluppi futuri. In relazione a ciò, i ricercatori intendono chiarire che, secondo la loro visione, hanno trattato l’aspetto probabilmente più ampio di questo campo; così, il contributo è ben lontano dall’essere un tentativo di riesaminare i progressi nella “Fisiologia della Psico-neuroimmunologia”. Sono enfatizzati alcuni aspetti dell’Immunofisiologia, come per esempio le interazioni attive tra il sistema immunitario (IS) e l’asse ipotalamo-ipofisi-surrene (HPA) o il sistema nervoso simpatico (SNS), e il ruolo delle citochine di origine periferica e centrale come mediatrici di queste interazioni. Le interazioni con altri sistemi endocrini e autonomi non sono menzionate, o lo sono solo marginalmente. Aspetti come stress, sonno e comportamento vengono considerati in altri articoli di questa serie.
Parte di questo articolo è stata scritta come una specie di “autobiografia scientifica” dato che, appartenendo alla “vecchia” generazione, è stato ritenuto dagli autori che condividere l’esperienza maturatapotrebbe essere interessante per le nuove generazioni che sicuramente saranno autrici delle svolte che questo campo necessita. A causa dello spazio limitato, il lavoro di molti colleghi non è stato citato e neanche accennato. Con qualche eccezione, le citazioni a partire da prima del 1987 e il 1996 sono state omesse. Queste citazioni si possono trovare in riviste meno recenti (Besedovsky and del Rey,1996; Besedovsky and Sorkin, 1997).
Inoltre, i ricercatori vogliono chiarire un aspetto che costituisce l’”essenza” della Fisiologia. Non c’è dubbio che il sistema immunitario è un sistema fisiologico omeostatico che, entro certi limiti, contribuisce alla stabilità e all’integrità dell’organismo (auto-conservazione e neutralizzazione dei pericoli). Comunque, causa di confusione potrebbe essere il fatto che, mentre le risposte immunitarie sono risposte fisiologiche che ci si aspetta efficienti al massimo durante le malattie infettivo–infiammatorie, esse talvolta contribuiscono ad innescare patologie. Così, sebbene il controllo e la regolazione dei diversi sistemi di un organismo sia al centro della Fisiologia, l’unica condizione, nel caso dell’immunoregolazione, è che tale processo fisiologico operi simultaneamente e collegato strettamente con gli eventi patologici. Un altro punto da notare è che la “Fisiologia” (come indica la sua etimologia derivante dal greco Physis = Natura) è una disciplina che tratta i processi naturali. Così, per esempio, ogni scoperta sull’effetto di un ormone su un dato parametro immunologico, seguendo un approccio farmacologico, dovrebbe essere seguita da studi che stabiliscano se tale effetto venga anche osservato sotto condizioni naturali, fisiologiche o patofisiologiche.
2. PRIMA DEL 1987: LA “VECCHIA” GENERAZIONE
Molti epidemiologi concordano sul fatto che l’acquisizione della conoscenza scientifica sia preceduta e suffragata da considerazioni deduttivo-intuitive. Il lavoro sulle interazioni immuno-neuro-endocrine 35 anni fa era in gran parte basato sull’intuizione poiché era fondato sulla convinzione che il sistema immunitario, come altri sistemi fisiologici, dovesse anche essere soggetto al controllo integrativo dei meccanismi neuro-endocrini. Infatti, i dati disponibili erano relativamente scarsi e basati, per esempio, sull’effetto della manipolazione di certe aree cerebrali e di meccanismi endocrini in alcuni processi immunitari che, in molti casi, non sono adattabili, come per esempio, nell’ipersensibilità acuta.
Recettori per solo pochi ormoni, come l’insulina, o neurotrasmettitori, come gli agenti adrenergici, sono stati identificati, o ipotizzati, sulle cellule immunitarie. C’è stato anche qualche studio che dimostrava delle interazioni tra i meccanismi neuro-endocrini e immunitari durante l’ontogenesi, come è il caso dell’effetto del timo sulla maturazione delle funzioni sessuali. Tuttavia, le informazioni disponibili sono state appena sufficienti per la formulazione iniziale dell’ipotesi che le risposte immunitarie sono soggette ad un livello di regolazione neuro-endocrina. Come succede con altre regolazioni fisiologiche, i meccanismi immunoregolatori neuro-endocrini devono essere basati sull’attività di scambi d’informazioni tra le cellule immunitarie e i sistemi nervoso ed endocrino. Poiché le risposte immunitarie, sia innate che acquisite, coinvolgono cellule e mediatori diversi a differenti stadi, la loro estrinseca regolazione dovrebbe essere basata su modificazioni neurali ed endocrine ben sincronizzate. Tali modificazioni, a loro volta, dovrebbero essere in grado di modulare l’attività delle cellule immunitarie su stadi definiti della risposta immunitaria.
Per verificare sperimentalmente l’ipotesi già menzionata, è stato necessario dimostrare che:
- le modificazioni neuro-endocrine che si verificano durante la risposta immunitaria non sono conseguenza della malattia o dello stress provocato dall’essere ammalato;
- le risposte neuro-endocrine immunologicamente indotte possono influenzare il funzionamento del sistema immunitario.
Seguendo questi criteri, è stato possibile dimostrare che i livelli di glucocorticoidi nel sangue sono aumentati in un modo soglia-dipendente durante il corso di specifiche risposte immunitarie ad antigeni innocui. E’ stato altresì dimostrato che l’aumento dei livelli di corticosterone durante la risposta ad un antigene può interferire con la risposta ad un secondo antigene non in relazione con il primo. Questa scoperta, che ha fornito una spiegazione al fenomeno della competizione tra antigeni, ha indicato che la modificazione endocrina osservata è stata rilevante per l’immunoregolazione. Sorprendentemente, mentre gli autori cercavano di indurre una competizione antigenica in vitro, hanno anche osservato che basse dosi di glucocorticoidi potevano aumentare il numero di cellule producenti specifici anticorpi.
La domanda che ci si è posto è stata: come possono scambiarsi informazioni due distinti sistemi del corpo, quello immunitario e quello endocrino? Per rispondere a questa domanda, sono state stimolate le cellule immunitarie con mitogeni o antigeni in vitro ed è stato scoperto che sovranatanti privi di cellule ottenuti da queste colture contenevano fattori capaci di stimolare l’asse ipotalamo-ipofisi-surrene (Besedovsky et al., 1981), un effetto che veniva mediato dall’ipofisi. Questa sostanza è stata chiamata “fattore che incrementa il glucocorticoide” (GIF). Fortunatamente, non ci si è sforzato eccessivamente di purificare questo fattore, perché, come si dirà dopo, varie citochine condividono la capacità di stimolare l’asse ipotalamo-ipofisi-surrene. Questa scoperta ha portato ad ipotizzare l’attività di un circuito immunoregolatore dell’asse immunitario ipotalamo-ipofisi-surrene.
Quasi nello stesso momento in cui tali studi furono pubblicati, Smith e colleghi riportarono che i glucocorticoidi interferiscono con la produzione del fattore di crescita della cellula T (come allora era chiamato IL-2) e degli altri prodotti immunitari. Venne anche dimostrato che i cambiamenti nei livelli endogeni di glucocorticoidi possono inibire la produzione di immunoglobulina. Inoltre, si scoprì che le rimanenti cellule immunitarie sono più sensibili agli effetti inibitori dei glucocorticoidi delle cellule attivate. Su queste basi, è stato ipotizzato che il circuito dell’asse immunitario ipotalamo-ipofisi-surrene può avere la funzione di prevenire l’eccessiva espansione di cellule a bassa affinità verso l’antigene e di quelle cellule reclutate sotto l’influenza policlonale delle linfochine. In questo modo, la specificità della risposta immunitaria potrebbe essere preservata e perfino migliorata. E’ intuibile che questo circuito, impedendo un’eccessiva espansione indistinta di cellule linfatiche e accessorie, gioca un suo ruolo nel prevenire malattie autoimmuni e linfoproliferative. Esempi sperimentali e clinici dimostranti l’importanza del circuito immunitario basato sull’asse ipotalamo-ipofisi-surrene saranno citati più avanti in quest’articolo.
| Figura 1. Il network immune-neuro-endocrino. I sistemi immunitario, endocrine e nervoso possono interagire a più livelli. Schematizzando, queste interazioni possono essere classificate come circuiti lunghi (linee blu) e brevi (linee verdi). Vi sono anche interazioni con il cervello (cerchio rosso). Le frecce che si diramano in marrone rappresentano altre conseguenze omeostatiche di tali interazioni. L’attività di queste interazioni può essere modificata dagli stimoli che agiscono principalmente sul sistema immunitario o sul cervello, per esempio stimoli psicosociali. Il risultato delle interazioni immuno-neuro-endocrine può comportare una modulazione immunitaria e/o comportamentale. |
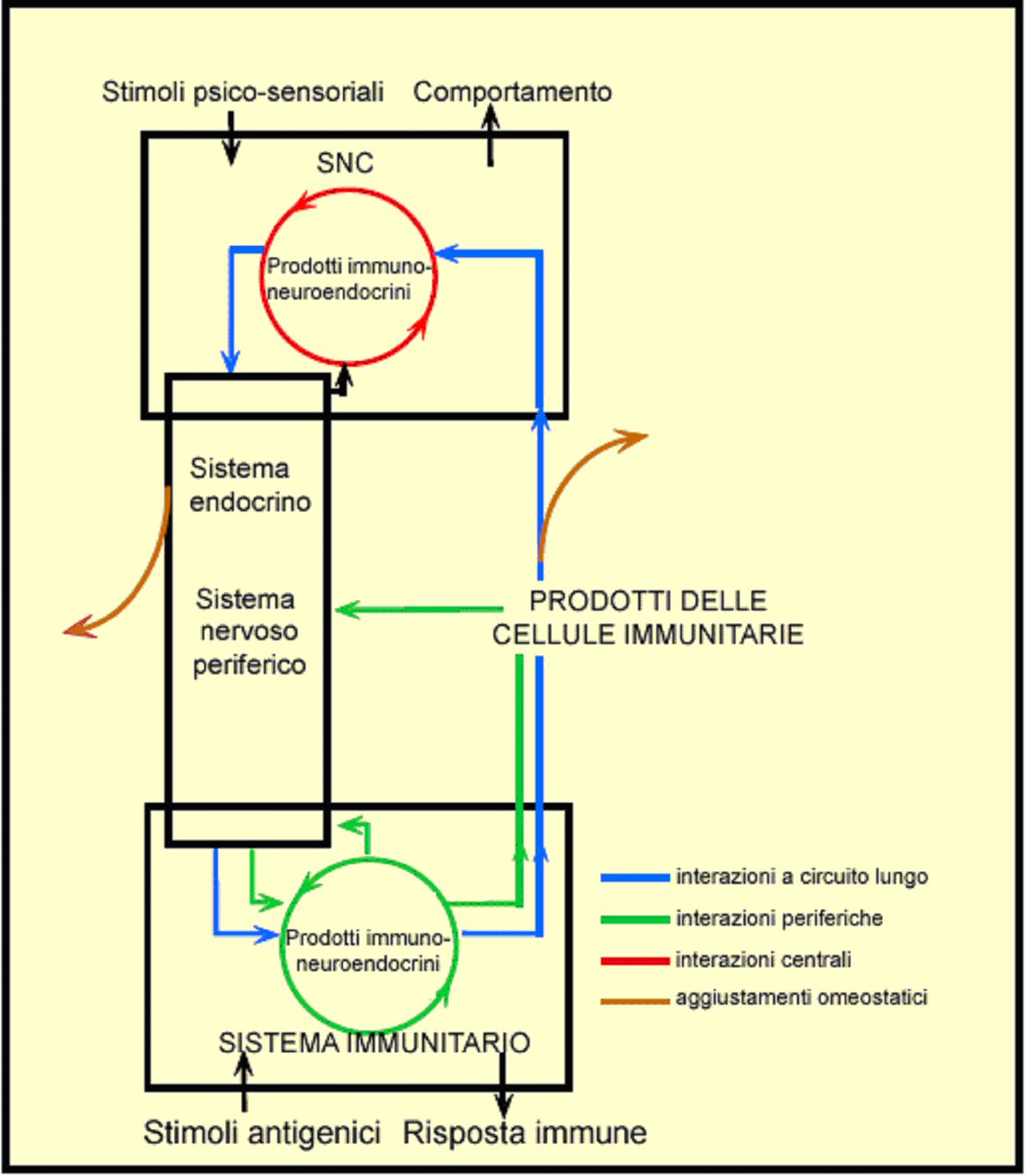 |
| L’ipotesi di un circuito basato sull’asse immunitario ipotalamo-ipofisi-surrene che controlla un’iper-produzione di prodotti immunitari è in linea con il concetto, formulato da A. Munck (che è stato il primo a caratterizzare il recettore per i glucocorticoidi), basato sull’affermazione che la funzione essenziale e generale dei glucocorticoidi è di controllare la reazione eccessiva di mediatori localmente prodotti. Intorno al 1986, divennero disponibili delle citochine in forma pura e ricombinante, rendendo possibile testare la loro capacità di indurre risposte neuro-endocrine. La IL-1 fu la prima citochina di cui si dimostrò la capacità di attivare l’asse ipotalamo-ipofisi-surrene (Besedovsky et al.,1986). L’interconnessione descritta, coinvolgendo meccanismi immunitari periferici e risposte endocrine sotto il controllo del cervello, portò al concetto dei circuiti immuno-neuro-endocrini “a ciclo lungo” (Fig. 1, linee azzurre) e fu sottoposta a un’intensa ricerca nelle seguenti decadi.
In molte condizioni, il sistema nervoso simpatico funziona in un modo coordinato con l’asse ipotalamo-ipofisi-surrene. Sembrava, quindi, logico studiare se esso partecipasse anche all’immunoregolazione. Su tale base, è stato indagato se la concentrazione di noradrenalina (NA) negli organi linfatici venga interessata durante la risposta immunitaria ad antigeni innocui. I risultati dimostrarono che una diminuzione nella concentrazione, nel contenuto e nel ricambio di noradrenalina (il principale Comunque, quest’interpretazione deve essere presa con cautela alla luce delle ulteriori scoperte (vedi sotto). Sebbene l’attività del sistema nervoso simpatico sia integrata centralmente, lo studio pionieristico di Felten e Bullock, dimostrante che le cellule immunitarie e le fibre nervose noradrenalinergiche sono in stretto contatto negli organi linfatici, hanno portato a ipotizzare l’esistenza di circuiti neurali immunoregolatori a ciclo breve (Fig.1, linee verdi). Negli anni ’80, si è iniziato a studiare anche la possibilità che la presenza di cellule neoplastiche, sia che vengano trapiantate, indotte de novo o in crescita spontanea, potesse indurre risposte endocrine. Chiare alterazioni endocrine, che avvenivano prima che il tumore diventasse palpabile e senza alcun segno visibile di malattia, si notarono durante la crescita del tumore. Le alterazioni coinvolgevano non solo gli ormoni E’ stato studiato, nello stesso animale, sia la risposta immunitaria che il tasso di attivazione di neuroni individuali in vari nuclei ipotalamici e a vari intervalli, dopo l’iniezione di antigeni innocui. Un evidente incremento del tasso di attivazione dei neuroni del nucleo ipotalamico ventromediale è stato rilevato parallelamente alla risposta a questo antigene. Questi risultati fornirono una prova diretta che il cervello riceve informazioni dal sistema immunitario in condizioni di assenza di stress nei ratti sotto narcosi, usando due diversi antigeni non-infettivi ed una metodologia convenzionale per valutare l’attività dei neuroni. E’ stato anche scoperto che il tasso di turnover della noradrenalina nei neuroni ipotalamici era inibito durante la risposta immunitaria verso eritrociti di pecora, un effetto che è verosimilmente mediato dai prodotti rilasciati dalle cellule immunitarie (Besedovsky et al., 1983). Lo studio dimostrante che la risposta immunitaria può, da sola, suscitare risposte neuro-endocrine e che i prodotti derivati dalle cellule immunitarie possono mediare tali risposte portò ad ipotizzare che il sistema immunitario agisce come un organo recettore periferico in grado di trasmettere informazioni al cervello riguardo a risposte a stimoli antigenici esterni o interni. Blalock e Smith scoprirono che le cellule immunitarie possono secernere “ormoni ipofisari” (Blalock et al., 1985). Inoltre, dato che le cellule cerebrali possono produrre citochine in passato descritte come prodotti immunitari, i due studiosi hanno evidenziato che c’era un uso comune di leganti e recettori dei mediatori immunitari e neuro-endocrini, e hanno chiamato il sistema immunitario “il sesto senso”. Questo periodo si è concluso con un validissimo contributo in questo campo: un simposio plenario sulle interazioni immuno-neuro-endocrine è stato organizzato per la prima volta in un congresso internazionale sull’Immunologia (VI Congresso Internazionale sull’Immunologia, luglio 1986, Canada). Sebbene questo simposio si sia svolto contemporaneamente ad un altro che trattava la “sensazionale” scoperta della restrizione dell’MHC, i “nostri” sono stati seguiti con entusiasmo da un pubblico numeroso. |
3. 1987-1996Un aumento quasi esplosivo del numero degli scienziati impegnati nella ricerca nel campo delle interazioni immuno-neuro-endocrine si è verificato nella decade 1987-1996. BBI non è solo stata pubblicata come conseguenza del vasto interesse in questo campo, ma ha anche contribuito alla sua crescita. La conoscenza del circuito dell’asse immunitario ipotalamo-ipofisi-surrene è stata approfondita da studi dimostranti che molte citochine condividono la capacità di stimolare tale asse. IL-1, IL-2, IL-3, IL-6, Il-8, IL-11, IL-12, TNF, INFGamma, e GM-CSF sono tra le citochine che possono integrare tale feedback mediato dal glucocorticoide. Il campo d‘azione delle citochine è stato un argomento di discussione. Ora è chiaro che l’effetto acuto delle citochine viene generalmente esercitato a livello dell’ipotalamo attraverso la secrezione del CRH e fino a un certo grado anche di AVP. Comunque, in condizioni prolungate, sono stati osservati anche effetti a livello dell’ipofisi e del surrene (vedi Besedovsky and del Rey, 1996; Turnbull and Rivier 1999). La consapevolezza dell’importanza del circuito di feedback dell’asse immunitario ipotalamo-ipofisi-surrene durante la malattia iniziò nel periodo che va dal 1987 al 1996, attraverso tre notevoli scoperte. Fu riportato da diversi gruppi che:
Si deve anche accennare che si è scoperto che, oltre all’asse ipotalamo-ipofisi-surrene, altri sistemi endocrini sono interessati nella risposta immunitaria (vedi Besedovsky and del Rey, 1996). I livelli cellulari in cui il SNS può esercitare delle azioni regolatrici sono stati ulteriormente caratterizzati in questo periodo (vedi Heijnen and Kavelaars, 1999). Le principali cellule bersaglio dei neurotrasmettitori adrenergici sembrano essere i timociti maturi e immaturi, le cellule timiche epiteliali, i linfociti T, i macrofagi, i mastociti, le cellule plasmatiche, e le cellule enterocromaffini. E’ stato evidenziato che la noradrenalina e l’adrenalina, stimolando la via di segnalamento recettore Beta2 adrenergico–AMP ciclico-proteina chinasi A, inibisce la produzione di citochine pro-infiammatorie tipo 1, per esempio IL-12, TNFAlfa e IFNGamma. La stimolazione dei recettori Beta-adrenergici inibisce le cellule presentanti antigeni e le cellule Th1, ma stimola la produzione di citochine anti-infiammatorie tipo 2 come IL-10 e TGFBeta. Così, le catecolammine endogene potrebbero causare una soppressone selettiva dell’infiammazione e dell’immunità cellulare Th1-mediate, favorendo in questo modo l’immunità umorale e proteggendo anche l’ospite dagli effetti nocivi delle citochine pro-infiammatorie e dagli altri prodotti dei macrofagi attivati (vedi Elenkov et al., 2000; Sanders, 2006). Si scoprirono anche effetti sinergici di glucocorticoidi e catecolammine (Elenkov et al., 2000). Ci furono pareri contrastanti sull’effetto della denervazione chirurgica del sistema simpatico e dello svuotamento chimico delle riserve di NA sulle risposte immunitarie. Gli autori non mostrano pregiudizi riguardo all’argomento perché hanno scoperto che entrambi i procedimenti danno luogo ad un aumento della risposta immunitaria agli eritrociti di pecora, un effetto che concorda con altri esperimenti riportati in cui sono stati usati antigeni diversi. Comunque, degli studi in cui è stata usata l’idrossi-dopamina 6 per svuotare le catecolammine hanno rivelato, in alcuni casi, effetti immunosoppressivi quando testati poco dopo la somministrazione della neurotossina. L’interpretazone è che il rilascio acuto di NA, causato dal danno del nervo terminale, e il marcato aumento dei livelli di corticosteroidi, indotto da tale processo, esercitano un effetto inibitorio sulle cellule immunitarie. E’ chiaro, comunque, che la denervazione sia un procedimento molto estremo il quale, pur rivelando effetti tonici da parte dei neuro-trasmettitori del sistema simpatico sull’attivazione, la differenziazione e la sopravvivenza della cellula immunitaria, non permette conclusioni definitive riguardo alle risposte immunitarie, le quali sono processi fasici,dinamici e stage-dipendenti. Per ulteriori informazioni sull’immunoregolazione del sistema simpatico ottenute nel periodo tra il 1987 e il 1997, si possono consultare le riviste Elenkov et al., 2000 e Sanders, 2006. Sebbene l’innervazione colinergica sia stata osservata nel timo, ma non negli organi linfatici periferici, come la milza, sono stati riferiti effetti dei mediatori para-simpatici sui processi immunitari. Per esempio, Rinner e Schauenstein hanno individuato un effetto inibitorio del carbacolo colinergico agonista su una risposta immunitaria agli eritrociti di pecora indotta in vitro. Si è scoperto anche che l’innervazione peptidergica integra i potenziali circuiti immunoregolatori a ciclo breve. Molti organi linfatici ricevono anche l’innervazione peptidergica sensoriale che si trova principalmente confinata al parenchima, e i peptidi maggiormente presenti sono tachichinine (sostanza P, neurochinina A), peptide calcitonina correlata al gene (CGRP), e polipeptide intestinale vasoattivo/peptide isoleucina istidina (VIP/PHI). Inoltre, si è provato che alcuni di questi peptidi co-esistono con le catecolammine e che esiste una stretta relazione spaziale tra fibre nervose peptidergiche e mastocisti, cellule T e macrofagi. Scarsi nervi peptidergici sembrano presenti nelle regioni della cellula B pura. Si osservano relativamente spesso contatti tra neuroni e mastocisti in tutti gli organi linfatici, a eccezione della milza (Weihe et al., 1991). Sebbene qualche prova è stata già fornita in precedenza, divenne ben consolidato tra il 1987 e il 1997 che le citochine immunitarie influenzano meccanismi complessi che coinvolgono una varietà di circuiti neuronali come la termoregolazione, l’assorbimento del cibo, gli schemi del sonno e il comportamento. Divenne chiaro che non esiste un singolo pirogeno endogeno dato che svariate sostanze endogene, come IL-1, IL-6, IL-8, IFNGamma, IFNBeta e GM-CSF possono provocare febbre. Anche varie citochine possono inibire l’assorbimento del cibo, tra cui IL-1, IL-6, IL-8 e TNFAlfa. La capacità di aumentare il sonno a onde lente viene anche condivisa da diverse citochine come IL-1, IL-2, IFNGamma, e TNFAlfa, un effetto che viene modulato da IL-4, IL-10 e IL-13. Si sa che varie citochine producono profondi effetti sul comportamento, per esempio sull’apprendimento, sulla capacità di esplorazione e sull’istinto di fuga. Pare che alcune di queste azioni avvengano a livello del sistema nervoso centrale e siano mediate da IL-1 dato che la somministrazione i.c.v. di antagonisti del recettore dell’IL-1 blocca tali effetti (Kent et al., 1992). Molti dei meccanismi integrati a livello cerebrale si trovano sotto il controllo dei neuroni cerebrali catecolaminergici e serotoninergici. Nello stesso momento in cui lo fece Dunn, abbiamo riferito che IL-1 stimola il tasso del turnover della noradrenalina nel cervello. Nel caso dei neuroni noradrenergici, l’effetto è stato non solo limitato al cervello, ma è stato anche riscontrato a livello del midollo spinale nel tratto inferiore, suggerendo una via metabolica neurale per gli effetti IL-1 nel sistema nervoso centrale. C’è stata anche la prova dell’importanza della stimolazione dei neuroni centrali noradrenergici per l’effetto di IL-1 sull’asse ipotalamo-ipofisi-surrene e sulla febbre (vedi Besedovsky e del Rey, 1996; Turnbull e Rivier, 1999). Un aspetto che sollevò grande interesse in questo periodo fu la comprensione del modo in cui i segnali immunitari possano raggiungere il cervello e influenzare le funzioni cerebrali. Lo studio dimostrò che sono coinvolte vie metaboliche umorali e neurali. Le vie metaboliche umorali possono mandare informazioni al cervello sia attraverso organi circumventricolari sia attraverso l’interfaccia endoteliale/gliale dei vasi sanguigni. I gruppi di Dantzer, Maier e Watkins, e Blatteis hanno dimostrato l’esistenza di una via metabolca neurale mettendo in luce il ruolo importante delle fibre afferenti vagali nella trasmissione dei segnali delle citochine al cervello (vedi Dantzer, 2004; Maier et al., 1998). In generale, i dati disponibili hanno indicato che la via metabolica umorale può venire seguita durante il processo immunitario che dà luogo ad alti livelli di citochine nella circolazione, mentre la via metabolica neurale sembra predominare quando tali mediatori vengono rilasciati localmente nei tessuti attraverso l’innervazione vagale. Inoltre, entrambe le vie possono funzionare simultaneamente oppure in sequenza durante le risposte immunitarie. Verso la fine del periodo che va dal 1987 al 1996, un evento importante per il campo delle interazioni tra il cervello e il sistema immunitario è stato la dimostrazione originariamente prodotta dai laboratori di Bartfai e Dantzer che l’attivazione delle cellule immunitarie periferiche mediante lipopolisaccaride induce una produzione di citochine nel cervello. Inizialmente, questi risultati preoccupavano perché la dose di lipopolisaccaride usata può distruggere la barriera emato-encefalica o causare alterazioni cerebrali come conseguenza dello shock endotossico. Comunque, quando sono stati fatti degli studi usando una dose di lipopolisaccaride che non intacca la barriera emato-encefalica e non causa evidenti sintomi di malattia, è stata individuata anche l’espressione genica della citochina nel cervello (Pitossi et al., 1997), confermando ed ampliando così i lavori precedenti. L’espressione genica per IL-1Beta, IL-6, TNFAlfa e IFNGamma viene aumentata attraverso una somministrazione periferica di lipopolisaccaride. L’inizio della trascrizione e il picco dell’accumulo di mRNA sono dipendenti sia dalla citochina che dalla regione cerebrale studiata. L’espressione di IL-1Beta e di IL-6 è generalmente aumentata nell’ipotalamo e nell’ippocampo, mentre l’espressione di TNFAlfa è più marcata nel talamo e nel nucleo striato. Nelle condizioni accennate, queste citochine sono meno inducibili nella corteccia cerebrale. Non sono state individuate correlazioni tra l’espressione genica della citochina e la densità delle strutture vascolari in una data area del cervello, né è stata osservata un’espressone di citochina preferenziale in aree del cervello che includono gli organi circumventricolari. Una funzione fisiologica principale del sistema nervoso simpatico è il controllo della pressione e del flusso del sangue nei tessuti. Questo processo è molto importante per l’efficacia della risposta immunitaria. Indubbiamente, le cellule immunitarie hanno bisogno di circolare per raggiungere i luoghi dove avvengono i processi infiammatori e infettivi. I sistemi circolatori sanguigno e linfatico, accoppiati, forniscono le vie per la circolazione delle cellule immunitarie, mentre le molecole di adesione, le chemochine, le integrine e altri mediatori localmente prodotti controllano la loro localizzazione e mobilizzazione. Inoltre, la milza è inserita nel sistema circolatorio in modo tale che favorisca la clearance, l’assorbimento e la ritenzione di microrganismi e il loro contatto con le cellule immunitarie. Nei roditori, quest’organo è privo di circolazione linfatica ed il controllo del flusso sanguigno è strettamente basato sul tono vascolare del sistema simpatico. Come è stato detto, è stato scoperto che una diminuzione dell’attività del sistema simpatico precede il picco di una specifica risposta immunitaria. Paradossalmente, si è dimostrato che la somministrazione di IL-1 e di lipopolisaccaride può incrementare l’attività del sistema nervoso simpatico. Quest ‘effetto non era compatibile con l’aumento del flusso sanguigno e con l’accumulo che era stato osservato negli organi linfatici durante il processo immunitario, il quale è probabilmente basato su una diminuzione del controllo del sistema simpatico sul tono vascolare. C’è comunque una spiegazione a queste scoperte apparentemente contraddittorie. La IL-1 prodotta negli organi linfatici aumenta selettivamente il flusso sanguigno interferendo localmente con il tono del simpatico, così provocando una redistribuzione del flusso sanguigno. Mentre la stimolazione del nervo splenico in vivo diminuisce il flusso sanguigno nella milza, questo effetto viene annullato completamente negli animali trattati con lipopolisaccaride o IL-1 come conseguenza di un’inibizione post-giunzionale del rilascio di noradrenalina. Alcuni studi in vitro hanno dimostrato che IL-6 e TNF provocano un effetto simile sul rilascio di noradrenalina da parte delle fibre nervose del sistema simpatico. Questi dati corroborano i dati discussi in precedenza sul fatto che le cellule immunitarie attivate siano meno esposte all’effetto delle fibre noradrenergici e fanno pensare ad un meccanismo attraverso il quale le citochine pro-infiammatorie potrebbero deviare l’afflusso di sangue verso i siti dove avviene il processo immunitario (Rogausch et al.1997). Questa ridistribuzione dell’afflusso del sangue favorisce il contatto tra cellule immunitarie e antigeni: un processo essenziale per una risposta immunitaria efficiente. II glucosio è la principale fonte di energia per il cervello e per la maggior parte dei tessuti periferici, incluse le cellule immunitarie. Inoltre, processi immunitari essenziali come l’endocitosi, la fagocitosi, l’aumentato turnover cellulare, l’espansione clonale, la produzione di numerosi mediatori, la generazione di cellule effettrici e di molecole sono molto dispendiosi in termini di energia. E’ stato dimostrato che basse, sub-pirogene dosi di IL-1 inducono nei topi un’ipoglicemia insulino-dipendente intensa e di lunga durata (del Rey ad Besedovsky, 1992). Quest’effetto, che viene anche osservato in animali insulino-resistenti, si sviluppa nei topi nonostante l’aumento dei livelli di ormoni controregolatori come le catecolammine, i glucocorticoidi e il glucagone. Ci sono anche dati che l’effetto ipoglicemico di IL-1 può essere indotto a livelli centrali dato che la somministrazione intra-cerebro-ventricolare delle citochine dà luogo a una riduzione dei livelli di glucosio nel sangue (del Reyet al,1998). Comunque, l’effetto più sorprendente viene osservato quando topi e ratti ricevono un challenge con carico di glucosio molte ore dopo una singola iniezione intra-peritoneale di IL-1 (vedi Besedovsky and del Rey, 1996). In tale situazione, si osserva chiaramente che, in seguito ad un’elevazione temporanea dei livelli di glucosio nel sangue, la sua concentrazione ritorna ai livelli ridotti precedenti e che gli animali a cui è stata praticata l’iniezione di IL-1 rimangono ipoglicemici per molte altre ore. Queste scoperte indicano chiaramente che l’IL-1 cambia il rigido settaggio che caratterizza l’omeostasi del glucosio. Questi cambiamenti sono legati alla capacità di indurre il trasporto e l’ossidazione del glucosio dimostrata da IL-1, per esempio nelle cellule adipose e nei fibroblasti; essi sono anche legati ad un effetto sul meccanismo gluco-regolatore sotto il controllo del cervello (del Rey and Besedovsky, 1992). La combinazione degli effetti locali e centrali dell’IL-1 servirebbe a deviare il glucosio verso gli organi linfatici e verso i tessuti infiammati/infetti per soddisfare l’alto dispendio di energia delle risposte immunitarie (Fig. 1 sopra, frecce color marrone). |
4. 1997-2006Dal 1997 in poi, la ricerca in questo campo è stata principalmente orientata a scoprire la rilevanza fisiologica e patologica delle interazioni immuno-neuro-endocrine ed a meglio comprendere la base molecolare di tali interazioni. L’importanza del circuito regolatore dell’asse immunitario ipotalamo-ipofisi-surrene nel controllo dei processi infiammatori ed autoimmuni è stata definitivamente stabilita (Besedovsky and del Rey, 2006; Sternberg, 2006). Considerevoli prove sperimentali e cliniche sottolineano l’importanza del circuito feedback dell’asse citochina-ipotalamo-ipofisi-surrene durante le malattie autoimmuni ed infettive. Per esempio, la surrenectomia o il blocco dei recettori dei glucocorticoidi aggravano la malattia e aumentano la mortalità in modelli animali di artrite reumatoide, sclerosi multipla, tiroidite di Hashimoto e shock settico. Una diminuita risposta dell’asse ipotalamo-ipofisi-surrene o una ridotta sensibilità ai glucocorticoidi sono connesse anche con patologie umane come malattie reumatiche, sclerosi multipla, sindrome di Sjogren, asma allergico, dermatite atopica, malattia infiammatoria intestinale e fibromialgia (vedi Besedovsky e del Rey, 2006; Sternberg, 2006). Le disfunzioni, notate in così tante patologie, vengono ora spiegate attraverso una migliore conoscenza delle azioni dei glucocorticoidi, le quali possono influenzare una grande quantità di mediatori infiammatori. Si deve aggiungere che molti effetti dei glucocorticoidi vengono modulati dall’ormone androgeno (DHEA) deidroepiandrosterone secreto dal surrene. In qualche caso, particolarmente nelle malattie croniche, il rapporto cortisolo/ DHEA riflette l’efficacia del circuito dell’asse citochina-ipotalamo-ipofisi-surrene. Questo circuito e la sua rilevanza in patofisiologia sono rappresentati schematicamente nella Fig. 2 come esempio perché, attualmente, questo è probabilmente il circuito immunoregolatore neuro-endocrino meglio descritto. |
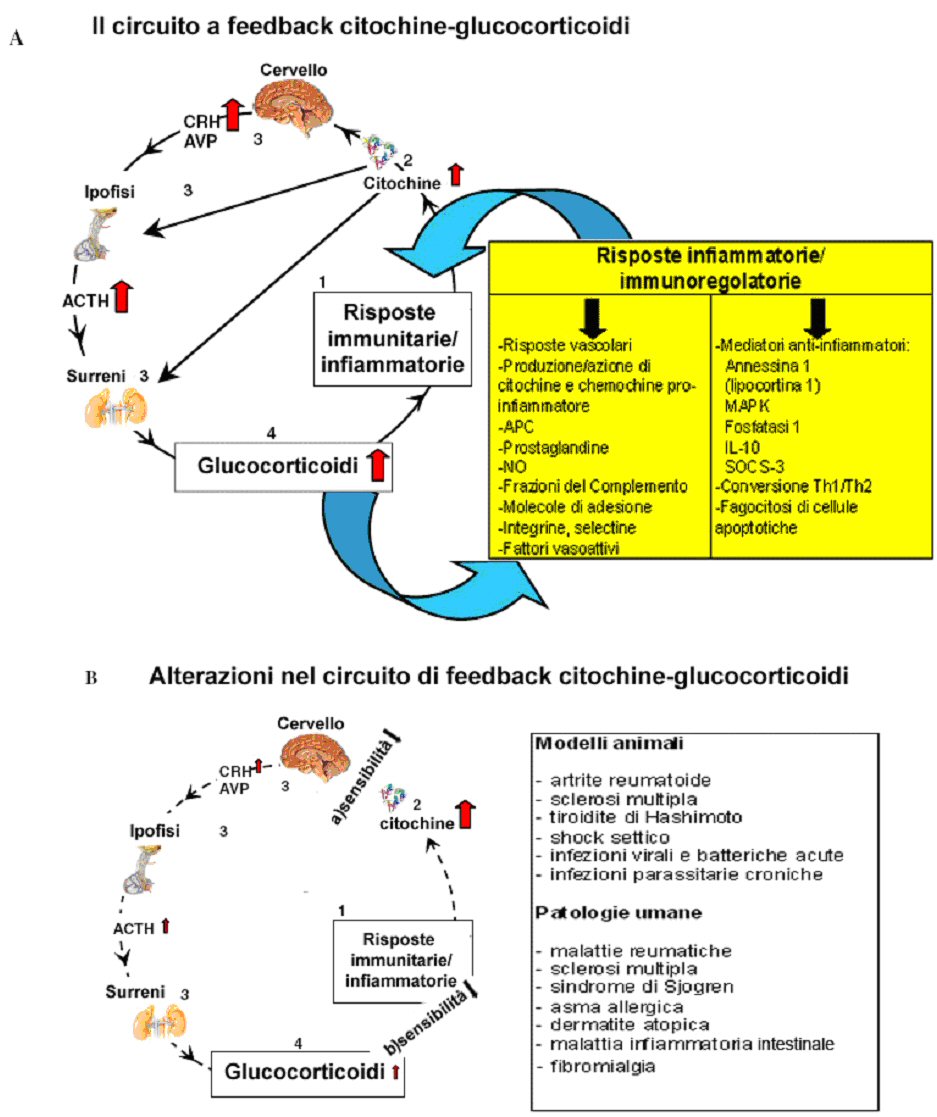 Figura 2. Il circuito a feedback dell’asse citochine-HPA. Questo circuito è dato come esempio di meccanismi immunoregolatori neuro-endocrini. (A) In seguito ad alcune risposte infiammatorie e immunitarie, le citochine, e in particolare quelle pro-infiammatorie, possono stimolare l’asse HPA a diversi livelli. Come conseguenza gli elevati livelli di glucocorticoidi possono modulare i processi infiammatori e immunitari stimolando o reprimendo diversi meccanismi o la produzione di mediatori. (B) Alterazioni a livello del circuito a feedback dell’asse citochine-HPA possono aggravare il decorso di alcune malattie. Dati sperimentali e clinici indicano che il circuito a feedback dell’asse citochine-HPA è alterato in patologie umane e in modelli animali di alcune malattie sia perché vi è una ridotta risposta dell’asse HPA alle citochine (a) o perché si sviluppa una resistenza ai glucocorticoidi (b). APC: cellule presentanti l’antigene; NO: ossido nitrico; MAPK: proteine chinasi attivate da mitogeni; SOCS: soppressori del segnalamento delle citochine. Figura 2. Il circuito a feedback dell’asse citochine-HPA. Questo circuito è dato come esempio di meccanismi immunoregolatori neuro-endocrini. (A) In seguito ad alcune risposte infiammatorie e immunitarie, le citochine, e in particolare quelle pro-infiammatorie, possono stimolare l’asse HPA a diversi livelli. Come conseguenza gli elevati livelli di glucocorticoidi possono modulare i processi infiammatori e immunitari stimolando o reprimendo diversi meccanismi o la produzione di mediatori. (B) Alterazioni a livello del circuito a feedback dell’asse citochine-HPA possono aggravare il decorso di alcune malattie. Dati sperimentali e clinici indicano che il circuito a feedback dell’asse citochine-HPA è alterato in patologie umane e in modelli animali di alcune malattie sia perché vi è una ridotta risposta dell’asse HPA alle citochine (a) o perché si sviluppa una resistenza ai glucocorticoidi (b). APC: cellule presentanti l’antigene; NO: ossido nitrico; MAPK: proteine chinasi attivate da mitogeni; SOCS: soppressori del segnalamento delle citochine. |
| Un aspetto innovativo legato all’immunoregolazione attraverso i nervi noradrenalinergici deriva dalla scoperta che la noradrenalina induce apoptosi nelle cellule linfatiche attraverso la stimolazione dei recettori Beta-adrenergici. Questa scoperta ha portato a chiedersi se gli effetti pro-apoptotici della noradrenalina possano influenzare le risposte immunitarie che danno luogo alla distruzione apoptosi-mediata di cellule T specifiche (per esempio la risposta immunitaria ai super-antigeni) oppure possano verificarsi nelle patologie immunitarie in cui l’apoptosi Fas-Fas ligando mediata è imperfetta. I super-antigeni, come l’enterotossina stafilococcica B (SEB), inducono una forte risposta proliferativaseguita dalla distruzione clonale di una porzione sostanziale di definite cellule T VBeta. Le cellule rimanenti mostrano non reattività in vitro. Abbiamo scoperto che, parallelamente alla risposta immunitaria all’enterotossina stafilococcica B si sono verificati cambiamenti bi-fasici nell’attività del sistema nervoso simpatico. Inoltre, la denervazione del simpatico ha dato origine a una diminuzione della proliferazione delle cellule SEB-indotte e ad una produzione di IL-2, e ha ostacolato la distruzione specifica delle cellule CD4VBeta4 della milza osservate in animali intatti senza interessare la non reattivià. Questi studi indicarono che l’effetto pro-apoptotico della noradrenalina può essere espresso in vivo e che è importante durante la risposta immunitaria ai super-antigeni. |
Quando sono stati studiati i meccanismi con cui la noradrenalina induce l’apoptosi delle cellule linfatiche, è stato scoperto che questo processo è indipendente da un Fas funzionale. Così, sono stati usati topi lpr/lpr, che sono privi dell’espressione del Fas funzionale (CD95) e sono, quindi, mancanti di un meccanismo critico per il mantenimento della tolleranza periferica, per indagare se i nervi noradrenergici possano influenzare l’espressione della malattia autoimmune linfoproliferativa che sviluppano spontaneamente.
Nella milza dei topi lpr/lpr, nei primi istanti dell’ontogenesi, la concentrazione di noradrenalina è significativamente aumentata, ma l’innervazione splenica del simpatico gradualmente diminuisce col progredire della malattia. Inoltre, i livelli sanguigni di IgM e la concentrazione splenica di noradrenalina sono inversamente correlati quando la malattia si manifesta in modo conclamato.
La simpatectomia nei neonati di topo, che anticipa sperimentalmente la perdita della denervazione che avviene spontaneamente nell’età adulta, dà luogo a una significativamente aumentata concentrazione di IgM e IgG2a nel sangue, accelera l’esordio di linfoadenopatie e accorcia notevolmente il tempo di sopravvivenza dei topi lpr/lpr. Questi dati dimostrano che, oltre alle imperfezioni nella via metabolica Fas, un’innervazione del sistema simpatico dei topi lpr/lpr contribuisce anche alla patogenesi della malattia autoimmune, e supporta fortemente l’ipotesi che il sistema nervoso simpatico possa modulare l’espressione delle malattie linfoproliferative (del Rey et l., 2002; del Rey e al., 2006).
Una valutazione generale delle opinioni odierne sull’immunoregolazione svolta dal simpatico può essere riassunta come segue. Le prove disponibili dimostrano che quasi tutti i meccanismi coinvolti in una risposta immunitaria possono essere influenzati dai neurotrasmettitori noradrenergici (vedi Elenkov et al., 2000; Sanders, 2006). Senza dubbio, la noradrenalina può inibire o stimolare una risposta immunitaria a seconda della dose di agonista somministrata e del tipo di recettore adrenergico stimolato. L‘effetto della noradrenalina dipende anche dal tipo di stimolo che innesca la risposta immunitaria, dal sottogruppo di cellule coinvolte, e, con maggior importanza, dalla fase della risposta durante la quale le cellule linfatiche e/o accessorie sono esposte ai neurotrasmettitori.
Tra i processi direttamente o indirettamente influenzati dai neurotrasmettitori del sistema simpatico ci sono la presentazione degli antigeni e l’espressione delle molecole co-stimolatorie e di adesione, l’attivazione delle cellule linfatiche, la produzione di citochine, l’espansione e il blocco clonale, la produzione di immunoglobuline, e la generazione di cellule citotossiche. Il fatto che la stimolazione simultanea dei recettori adrenergici e del recettore della cellula T influenzi vie comuni di segnalamento intra-cellulare potrebbe spiegare perché gli agonisti adrenergici possano influenzare una così grande quantità di meccanismi coinvolti in una risposta immunitaria.
L’interesse nella comprensione dell’importanza dell’altro ramo del sistema nervoso autonomo, il sistema para-simpatico, è aumentato negli ultimi anni. Esistono dei segnali che indicano che le fibre efferenti del nervo vago esercitano un ruolo protettivo nello shock endo-tossico. La prova è basata sul fatto che la vagotomia cervicale bilaterale aggrava l’abbassamento della pressione del sangue causata a una dose letale di LPS e sul fatto che la stimolazione delle fibre efferenti vagali modulano questo effetto. La conclusione che il nervo vago protegge l’ospite dallo shock endo-tossico è stata anche ricavata dal fatto che l’aumento di citochine, come il TNFAlfa, avviene negli animali vagotomizzati, mentre la stimolazione delle fibre efferenti del vago inverte quest’effetto.
Inoltre, usando un modello di infiammazione mediato dalla carragenina, si è dimostrato che l’attivazione dei recettori della nicotina, sia attraverso la stimolazione del nervo vago che attraverso gli agonisti colinergici, inibisce significativamente il rilascio di citochine pro-infiammatorie e blocca la migrazione dei leucociti. Esperimenti in vitro, in cui vennero usati macrofagi umani, hanno confermato che l’acetilcolina e gli antagonisti colinergici inibiscono il rilascio di TNF, IL-1 e IL-18 in risposta all’endo-tossina e che quest’effetto viene esercitato a livelli post-trascrizionali. Gli studi farmacologici indicano chiaramente che la stimolazione dei recettori colinergici della nicotina possono essere coinvolti nel controllo delle risposte infiammatorie esagerate (vedi Tracey, 2002). Questi studi, e quelli precedenti, costituiscono un aspetto importante della ricerca sulle interazioni immuno-neuro-endocrine che più probabilmente dimostra la rilevanza del sistema nervoso para-simpatico per l’immunoregolazione anche in condizioni meno estreme dello shock endo-tossico.
L’importanza del rilascio del neuro-peptide durante i processi infiammatori autoimmuni è diventata più evidente e quest’argomento è stato esaminato da varie riviste. Alcuni neuro-peptidi, in particolare SP, sono chiaramente pro-infiammatori, mentre altri, come VIP e PACAP, sono anti-infiammatori. Questi effetti sono stati riscontrati in modelli diversi di patologie, come lo shock settico, l’artrite reumatoide e la sclerosi multipla (Delgado et al., 2004; Jessop, 2002). Bisogna ancora chiarire fino a che punto la stimolazione dell’asse ipotalamo-ipofisi-surrene che risulta dalla somministrazione di questi neuro-peptidi contribuisca a tali effetti (Nussdorfer and Malendowicz, 1998).
Esistono oggi prove considerevoli che le citochine originate nel cervello possono influenzare i meccanismi del sistema nervoso centrale. Si è dimostrato che le citochine sono “fattori del sonno” e che influenzano sia i movimenti oculari non-rapidi che la fase di sonno REM. Livelli cerebrali di IL-1 e TNF concorrono alla propensione al sonno; per esempio, il loro livello aumenta dopo la privazione del sonno. Inoltre, la neutralizzazione immunitaria di IL-1 o il blocco del suo recettore nel cervello influenza il sonno a onde lente (Obal and Krueger 2003), indicando che IL-1 e TNF endogeni sono parte di una complessa cascata biochimica che regola il sonno. E’ stato anche dimostrato che l‘espressione di IL-1 può essere indotta nel cervello in stato di stress (vedi Besedovsky and del Rey, 1996) e che alcuni dei sintomi di “sickness behavior” sono integrati dalle citochine prodotte nel cervello. Per “sickness behavior” si intende un insieme coordinato di cambiamenti soggettivi, comportamentali e psicologici che si sviluppano negli individui malati nel corso di infezioni acute. Questi cambiamenti sono causati dagli effetti di IL-1 e di altre citochine pro-infiammatorie su bersagli cellulari del cervello. Senza dubbio, l’interferenza con gli effetti di queste citochine nel cervello impedisce l’espressione di certi sintomi del “sickness behavior”. Questi dati indicano il ruolo delle citochine pro-infiammatorie nel generare un “sickness behavior” durante patologie acute (Dantzer, 2004).
Lo studio del possibile ruolo delle citochine, prodotte nel sistema nervoso centrale, nella fisiologia del cervello e del loro contributo all’integrazione delle interazioni tra sistema immunitario e cervello a livello centrale è stato affrontato negli ultimi anni. Ci si è chiesto soprattutto se un aumento dell’attività neuronale in un cervello “sano” potesse influenzare la produzione locale di citochine. Una prova diretta deriva dalla dimostrazione che la stimolazione pre-sinaptica di definiti neuroni, come accade in condizioni fisiologiche, può controllare la produzione locale di citochine da parte delle cellule gliali e dei neuroni. Il potenziamento a lungo termine (LTP) dell’attività sinaptica nell’ippocampo è servita come modello per affrontare quest’argomento. Un chiaro aumento dell’espressione genica di IL-1Beta, innescata da neuroni glutaminergici attraverso recettori NMDA, è stato osservato in sezioni di ippocampo e in ratti liberi di muoversi nel corso di LTP (Schneider et al. 1998). Più recentemente, abbiamo osservato che il gene IL-6 è anche over-espresso in corso di LTP in vivo e in vitro (Balschun et al., 2004). Questi dati costituiscono la prima prova che l’espressione genica della citochina nel cervello può essere innescata da un indotto aumento pre-sinaptico dell’attività in una popolazione specifica di neuroni.
Gli autori dell’articolo e altri hanno studiato fino a che punto le citochine prodotte nel cervello durante LTP possano influenzare la plasticità e la performance delle sinapsi. A questo stadio, è necessario distinguere tra studi basati su una somministrazione esogena di citochine e studi focalizzati sugli effetti delle citochine prodotte endogenamente dalle cellule cerebrali. Esiste una vasta letteratura dimostrante che la somministrazione esogena di citochine in vivo ed in vitro può influenzare l’induzione di LTP e la plasticità delle sinapsi (vedi Besedovsky and del Rey, 1996). Questi studi, sebbene importanti dal punto di vista farmacologico, non possono dimostrare con affidabilità l’effetto delle citochine prodotte nel cervello in condizioni naturali. Infatti, il potenziamento a lungo termine è un fenomeno complesso che coinvolge un numero di recettori e mediatori che influenzano la sua inducibilità, il suo avviamento e il suo mantenimento in modi diversi.
In particolare, il mantenimento di LTP è dipendente dalla sintesi proteica e implica l’attivazione di geni in una data sequenza ed il rilascio dei loro prodotti in una certa quantità. Così, è quasi impossibile mimare l’effetto tempo-dipendente di una citochina prodotta endogenamente attraverso la sua somministrazione esogena. Per esempio, come sarà discusso in seguito, l’IL-1, una citochina che inibisce il potenziamento a lungo termine quando è somministrata esogenamente, contribuisce al consolidamento e al mantenimento di questo processo quando esso viene prodotto endogenamente. Usando lo specifico antagonista del recettore dell’IL-1 (IL-1ra), è stato scoperto che il blocco dei recettori IL-1, sia in vivo che in sezioni di ippocampo, dà luogo all’inibizione del mantenimento di LTP.
Quest’effetto è reversibile e avviene solo quando l’antagonista viene somministrato dopo che il potenziamento è innescato, cioè in un momento in cui, secondo gli studi già menzionati, ci si aspettano aumentati livelli di IL-1. Studi nei topi knock out per il recettore di tipo 1 dell’IL-1 sono in linea con queste scoperte (Avital et al., 2003). E’ stato recentemente scoperto che, in contrasto con l’effetto di supporto di IL-1, IL-6 contribuisce alla cessazione di LTP ben consolidato (Balschun et al., 2004). Nel complesso, questi risultati suggeriscono fortemente che IL-1Beta e IL-6 possono controllare il mantenimento dell’LTP nel cervello, un processo a cui è assegnato un ruolo nella formazione della memoria e in alcuni tipi di apprendimento. Inoltre, questi studi forniscono la prova del ruolo fisiologico e neuro-modulatore delle citochine originalmente descritte come mediatori immunitari.
Come nel caso del LTP, gli effetti della somministrazione di citochine sull’apprendimento, la memoria e il comportamento in generale, sono stati ampiamente studiati (vedi Anisman et al., 2005). Ancora una volta, questi studi sono indubbiamente di rilevanza farmacologica, ma non possono rispecchiare gli effetti fisiologici delle citochine, cosa che è il principale obiettivo di questo articolo. Così, di seguito saranno discussi i possibili effetti fisiologici delle citochine endogene sulla memoria e l’apprendimento. Come detto prima, un temporaneo blocco di IL-1 endogena nelle sezioni di ippocampo e nel cervello di ratti liberi di muoversi dà luogo all’inibizione del mantenimento del processo LTP. Considerando che è attualmente accettato che il processo LTP sia alla base di certe forme di memoria, è stato ipotizzato che tale processo sarebbe inibito in animali in cui gli effetti di IL-1 non possono essere visibili.
Questo è il caso dei topi knock out per il recettore IL-1 di tipo 1 (Yirmiya et al., 2002). Questi topi mostrano una più lunga fase di latenza a raggiungere una piattaforma nascosta, nella versione spaziale del test del labirinto ad acqua inoltre, mostrano una paura contestuale condizionata diminuita, ma si comportano allo stesso modo degli animali di controllo nei compiti di memoria ippocampo-indipendente. Il blocco dei recettori dell’IL-1 nel cervello di normali animali che eseguono un compito di apprendimento (il labirinto ad acqua di Morris) causa un handicap nella memoria ippocampo-dipendente. Questi risultati suggeriscono che il segnalamento dell’IL-1 all’interno dell’ippocampo gioca un ruolo critico nei processi di apprendimento e di memoria (Avital et al., 2003). E’ degno di nota il fatto che, negli studi precedentemente citati, il blocco del segnalamento di IL-1 è stato indotto dopo la procedura di training. D’altra parte, c’è la prova che il blocco prima del training degli effetti dell’IL-1 mediato da un vettore adenovirus esprimente l’antagonista IL-1ra, causa un miglioramento dei punteggi di conservazione della memoria sia a breve che a lungo termine (Depino et al., 2004). Comunque, come già detto, la IL-1 prodotta endogenamente nell’apprendimento contribuisce a memorizzare un preciso compito.
E’ stato anche studiato il ruolo dell’IL-6 endogenamente prodotta nel cervello. Come è già stato discusso, l’IL-6 viene prodotta durante il processo LTP. Il blocco di IL-6 endogena dopo l’apprendimento con alternanza spaziale ippocampo-dipendente ha dato luogo a significativi miglioramenti della memoria a lungo termine (Balschun et al., 2004). Inoltre, i topi knock-out per l’IL-6, nell’apprendimento al labirinto a raggi, si sono dimostrati facilitati per oltre 30 giorni per quanto riguarda un numero più basso di errori della memoria di lavoro (Braida et al., 2004).
Considerando questi risultati nel complesso, sembra chiaro che, pur avendo ruoli opposti, l’IL-1 e l’IL-6 endogene, prodotte nel cervello “sano”, sono importanti per il controllo della plasticità delle sinapsi e del processing della memoria nell’ippocampo. Il meccanismo che è alla base del ruolo dell’IL-1 e l’IL-6 in questi processi è ancora sconosciuto, ma dati recenti indicano il coinvolgimento di NFk-B, un fattore di trascrizione che media la produzione e gli effetti di citochine multiple (vedi Meffert and Baltimore, 2005).
Concludendo, esistono esempi che le citochine prodotte in un cervello in buone condizioni possano contribuire alla fisiologia cerebrale controllando l’attività dei neuroni, influenzando così i sistemi di controllo neuro-endocrini, la messa a punto della regolazione dei meccanismi omeostatici essenziali e le funzioni intrinseche del sistema nervoso centrale, come la memoria e l’apprendimento. C’è anche la prova che le citochine prodotte nel cervello coordinano gli adattamenti fisiologici durante le malattie e le risposte immunoregolatrici neuro-endocrine (Fig. 1, cerchio rosso). Sembra che le citochine giochino un doppio ruolo nelle patologie del cervello; da una parte esse possono controllare la risposta immunitaria locale attraverso interazioni citochine-neuroni e dall’altra si possono comportare come mediatori effettori in grado di contribuire alla patologia.
5. 2007: ASPETTANDO L’INIZIO
Questo sottotitolo è chiaramente ottimistico, dato che ha la presunzione di sottolineare la mole quasi infinita di lavoro lasciato alle nuove generazioni. Per esempio, spesso si è fatto riferimento alle risposte immunoregolatrici neuro-endocrine che vengono suscitate seguendo la stimolazione del sistema immunitario. Questa è un’eccessiva semplificazione che deriva dai modelli sperimentali usati inizialmente per portare alla luce delle interazioni immuno-neuro-endocrine. Non esiste “una” risposta immunitaria: ci sono probabilmente tanti tipi di risposte immunitarie quante sono le condizioni patologiche.
Mentre gli immunologi distinguono fra gli aspetti differenti dell’immunità naturale e il tipo della risposta immunitaria adattata, per esempio le risposteTh1/Th2-mediate, nelle condizioni reali l’immunità è basata su meccanismi incrociati che in molti casi sono il prodotto di un bilanciamento tra le cellule immunitarie e l’agente causale di una malattia, per esempio un microrganismo, e, perché no, l’intervento terapeutico usato. In ogni stadio di una malattia, vengono attivati meccanismi mediati da modelli molecolari patogeno-associati (PAMPs), vengono scoperte combinazioni particolari di cellule immunitarie e dei loro specifici prodotti e viene individuato un cocktail di altri mediatori nella circolazione e nei tessuti infiammati/infetti. Così, ci si aspetta che le risposte neuro-endocrine durante la malattia ed il loro risultato immunoregolatore siano differenti e che richiedano un esame a tutti i livelli. Inoltre oggi potremmo stabilire che le interazioni locali immuno-neuro-endocrine a ciclo breve potrebbero verosimilmente avvenire in tutti gli organi o tessuti sia in condizioni di salute che di malattia.
Nell’ultima decade, una svolta importante è derivata dall’esplorazione del complesso macchinario intracellulare implicato nella risposta di una cellula in seguito alla stimolazione di un dato recettore. Per esempio, le differenti proteine-chinasi, i secondi messaggeri, i fattori di trascrizione, gli eventi post-trascrizionali e i modulatori intra-cellulari possono diventare punti d’incontro durante l’attivazione simultanea o sequenziale dei recettori per mezzo di leganti di derivazione immunitaria e neuro-endocrina su cellule immunitarie e non-immunitarie. Questo darebbe luogo ad un potenziale sinergismo, o antagonismo, quasi infinito tra gli effetti di questi leganti in condizioni normali o patologiche. Comunque, dal punto di vista della Fisiologia, l’esame delle interazioni immuno-neuro-endocrine a livello cellulare non dovrebbe essere disgiunto dagli eventi omeostatici a monte che danno luogo a cambiamenti nella produzione e nella liberazione di diversi leganti ed all’espressione dei loro recettori sulla cellula bersaglio.
Oggi non suonerebbe esagerato affermare che quasi tutte le patologie hanno una componente infiammatoria/immunitaria che interagisce con i meccanismi nervosi ed endocrini. Infatti, le cellule immunitarie, in particolare le cellule dotate di antigeni, sono presenti in tutti i tessuti dove esse sono esposte ad ormoni e neuro-trasmettitori. Come è stato affermato nell’introduzione, è difficile trovare il limite tra situazioni fisiologiche e patologiche quando ci si riferisce al sistema immunitario. Gli autori si domandano se le diverse denominazioni date al campo siano in qualche modo restrizioni auto-imposte.
Per esempio, un campo di ricerca dedito allo studio della psico-neuro-immunologia delle malattie della pelle dovrebbe chiamarsi psico-neuro-immuno-dermatologia? Questo si dovrebbe applicare a tutte le specialità mediche ed anche alla Biologia (psico-neuro-immuno-biologia?) e si potrebbe continuare aggiungendo tutti i possibili acronimi della denominazione del nostro campo. Cosa stiamo realmente facendo quando studiamo le interazioni immuno-neuro-endocrine in ogni organo? Dal punto di vista degli autori, ciò che affrontano è un serio tentativo di integrare l’attuale conoscenza in biologia e in medicina, basata su un approccio multidisciplinare.
Questo tentativo serve a unire i dati ottenuti usando approcci sistemici con i dati derivati dalla realizzazione di tecnologie raffinate che permettono l’analisi di fenomeni a livello molecolare. Il lavoro presentato serve a ristabilire l’equilibrio tra deduzione e induzione, analisi e sintesi, cosa che è, in effetti, l’essenza della scienza. Il bisogno di ristabilire tale equilibrio deriva dall’attuale predominanza degli approcci analitici che talvolta degenerano in punti di vista riduttivi. Tale predominanza deriva probabilmente dagli immensi progressi tecnologici delle ultime decadi che danno luogo alla tentazione (talvolta favorita da enti di finanziamento e da giornali ad alto indice d’impatto) di investire risorse nello studio di meccanismi molecolari isolati dal loro significato fisiologico e pato-fisiologico. Senza dubbio, anche se tutti i prodotti che servono come messaggeri per la comunicazione intra-, extra- e inter-cellulare fossero identificati, non saremmo capaci di decifrare tutti i messaggi che essi potrebbero trasmettere. Un messaggero può essere purificato, ma non il messaggio che esso trasmette, messaggio che dipende da una grande varietà di condizioni spaziali e temporali e dalle condizioni attuali del bersaglio che riceve le informazioni. Così, la domanda è: è arrivato il momento per ri-denominare il nostro campo chiamandolo semplicemente “Biologia e Medicina Integrative”?
6. NOTE BIBLIOGRAFICHE
Ader, R., Cohen, N., 1975. Behaviorally conditioned immunosuppression. Psychosom. Med. 37, 333–340.
Anisman, H., Merali, Z., Poulter, M.O., Hayley, S., 2005. Cytokines as a precipitant of depressive illness: animal and human studies. Curr. Pharm. Des. 11, 963–972.
Avital, A., Goshen, I., Kamsler, A., Segal, M., Iverfeldt, K., Richter- Levin, G., Yirmiya, R., 2003. Impaired interleukin-1 signaling is associated with deficits in hippocampal memory processes and neural plasticity. Hippocampus 13, 826–834.
Balschun, D., Wetzel, W., Del Rey, A., Pitossi, F., Schneider, H., Zuschratter, W., Besedovsky, H.O., 2004. Interleukin-6: a cytokine to forget. FASEB J. 18, 1788–1790.
Besedovsky, H.O., del Rey, A., 1996. Immune–neuro–endocrine interactions: facts and hypotheses. Endocr. Rev. 17, 64–102.
Besedovsky, H.O., del Rey, A., 2006. Regulating inflammation by glucocorticoids. Nat. Immunol. 7, 537.
Besedovsky, H.O., del Rey, A., Sorkin, E., 1981. Lymphokine-containing supernatants from con A-stimulated cells increase corticosterone blood
levels. J. Immunol. 126, 385–387.
Besedovsky, H.O., del Rey, A., Sorkin, E., Da Prada, M., Burri, R., Honegger, C., 1983. The immune response evokes changes in brain noradrenergic neurons. Science 221, 564–566.
Besedovsky, H.O., del Rey, A., Sorkin, E., Dinarello, C.A., 1986. Immunoregulatory feedback between interleukin-1 and glucocorticoid hormones. Science 233, 652–654.
Besedovsky, H.O., Sorkin, E., 1977. Network of immune-neuroendocrine interactions. Clin. Exp. Immunol. 27, 1–12.
Blalock, J.E., Harbour-McMenamin, D., Smith, E.M., 1985. Peptide hormones shared by the neuroendocrine and immunologic systems. J. Immunol. 135, 858s–861s.
Braida, D., Sacerdote, P., Panerai, A.E., Bianchi, M., Aloisi, A.M., Iosue, S., Sala, M., 2004. Cognitive function in young and adult IL (interleukin)-6 deficient mice. Behav. Brain. Res. 153, 423–429.
Dantzer, R., 2004. Cytokine-induced sickness behaviour: a neuroimmune response to activation of innate immunity. Eur. J. Pharmacol. 500, 399–411.
del Rey, A., Besedovsky, H., 1989. Antidiabetic effects of interleukin 1. Proc. Natl. Acad. Sci. USA 86, 5943–5947.
del Rey, A., Besedovsky, H.O., 1992. Metabolic and neuroendocrine effects of pro-inflammatory cytokines. Eur. J. Clin. Invest. 22 (Suppl. 1), 10–15.
del Rey, A., Kabiersch, A., Petzoldt, S., Besedovsky, H.O., 2002. Involvement of noradrenergic nerves in the activation and clonal deletion of T cells stimulated by superantigen in vivo. J. Neuroimmunol. 127, 44–53.
del Rey, A., Monge-Arditi, G., Besedovsky, H.O., 1998. Central and peripheral mechanisms contribute to the hypoglycemia induced by interleukin-1. Ann. N. Y. Acad. Sci. 840, 153–161.
del Rey, A., Roggero, E., Kabiersch, A., Schafer, M., Besedovsky, H.O., 2006. The role of noradrenergic nerves in the development of the lymphoproliferative disease in Fas-deficient, lpr/lpr mice. J. Immunol. 176, 7079–7086.
Delgado, M., Pozo, D., Ganea, D., 2004. The significance of vasoactive intestinal peptide in immunomodulation. Pharmacol. Rev. 56, 249–290.
Depino, A.M., Alonso, M., Ferrari, C., del Rey, A., Anthony, D., Besedovsky, H., Medina, J.H., Pitossi, F., 2004. Learning modulation by endogenous hippocampal IL-1: blockade of endogenous IL-1 facilitates memory formation. Hippocampus 14, 526–535.
Elenkov, I.J., Wilder, R.L., Chrousos, G.P., Vizi, E.S., 2000. The sympathetic nerve—an integrative interface between two supersystems: the brain and the immune system. Pharmacol. Rev. 52, 595–638.
Heijnen, C.J., Kavelaars, A., 1999. The importance of being receptive. J. Neuroimmunol. 100, 197–202.
Jessop, D.S., 2002. Neuropeptides: modulators of immune responses in health and disease. Int. Rev. Neurobiol. 52, 67–91.
Kent, S., Bluthe, R.M., Dantzer, R., Hardwick, A.J., Kelley, K.W., Rothwell, N.J., Vannice, J.L., 1992. Different receptor mechanisms mediate the pyrogenic and behavioral effects of interleukin 1. Proc. Natl. Acad. Sci. USA 89, 9117–9120.
Maier, S.F., Goehler, L.E., Fleshner, M., Watkins, L.R., 1998. The role of the vagus nerve in cytokine-to-brain communication. Ann. N. Y. Acad. Sci. 840, 289–300.
Meffert, M.K., Baltimore, D., 2005. Physiological functions for brain NFkappaB. Trends Neurosci. 28, 37–43.
Nussdorfer, G.G., Malendowicz, L.K., 1998. Role of VIP, PACAP, and related peptides in the regulation of the hypothalamo–pituitary– adrenal axis. Peptides 19, 1443–1467.
Obal Jr., F., Krueger, J.M., 2003. Biochemical regulation of non-rapideye- movement sleep. Front. Biosci. 8, d520–d550.
Pitossi, F., del Rey, A., Kabiersch, A., Besedovsky, H., 1997. Induction of cytokine transcripts in the central nervous system and pituitary following peripheral administration of endotoxin to mice. J. Neurosci. Res. 48, 287–298.
Rogausch, H., del Rey, A., Kabiersch, A., Reschke, W., Ortel, J., Besedovsky, H., 1997. Endotoxin impedes vasoconstriction in the spleen: role of endogenous interleukin-1 and sympathetic innervation. Am. J. Physiol. 272, R2048–R2054.
Sanders, V.M., 2006. Interdisciplinary research: noradrenergic regulation of adaptive immunity. Brain Behav. Immun. 20, 1–8.
Schauenstein, K., Fassler, R., Dietrich, H., Schwarz, S., Kromer, G., Wick, G., 1987. Disturbed immune–endocrine communication in autoimmune disease. Lack of corticosterone response to immune signals in obese strain chickens with spontaneous autoimmune thyroiditis. J. Immunol. 139, 1830–1833.
Schneider, H., Pitossi, F., Balschun, D., Wagner, A., del Rey, A., Besedovsky, H.O., 1998. A neuromodulatory role of interleukin- 1beta in the hippocampus. Proc. Natl. Acad. Sci. USA 95, 7778– 7783.
Sternberg, E.M., 2006. Neural regulation of innate immunity: a coordinated nonspecific host response to pathogens. Nat. Rev. Immunol. 6, 318–328.
Sternberg, E.M., Hill, J.M., Chrousos, G.P., Kamilaris, T., Listwak, S.J., Gold, P.W., Wilder, R.L., 1989. Inflammatory mediator-induced hypothalamic–pituitary–adrenal axis activation is defective in streptococcal cell wall arthritis-susceptible Lewis rats. Proc. Natl. Acad. Sci.USA 86, 2374–2378.
Tracey, K.J., 2002. The inflammatory reflex. Nature 420, 853–859.
Turnbull, A.V., Rivier, C.L., 1999. Regulation of the hypothalamic– pituitary–adrenal axis by cytokines: actions and mechanisms of action. Physiol. Rev. 79, 1–71.
Weihe, E., Nohr, D., Michel, S., Muller, S., Zentel, H.J., Fink, T., Krekel, J., 1991. Molecular anatomy of the neuro–immune connection. Int. J. Neurosci. 59, 1–23.
Yirmiya, R., Winocur, G., Goshen, I., 2002. Brain interleukin-1 is involved in spatial memory and passive avoidance conditioning. Neurobiol. Learn Mem. 78, 379–389.